ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Цикл трикарбонових кислот (цикл Кребса)
У ході аеробного окиснювання вуглеводів піровиноградна кислота піддається окисному декарбоксилюванню з утворенням ацетил-КоА.
Ацетил-КоА, що утворився, вступає в різні реакції обміну й окислюється до СО2 і Н2О. Повне окиснювання ацетил-КоА відбувається в циклі трикарбонових кислот - циклі Кребса.
70 % ПВК окисляється до СО2 і Н2О через стадію утворення ацетил-КоА. При обміні білків, жирів і вуглеводів протягом доби на кожен 1 кг маси тіла утворюється близько 10 г ацетату.
Цикл Кребса є центром, де сходяться всі метаболічні шляхи, він - загальний кінцевий шлях окиснювання ацетильних груп (у вигляді ацетил-КоА), у які перетворюється в процесі катаболізму більша частина органічних молекул, що грають роль "клітинного палива" - вуглеводів, жирних кислот, амінокислот.
Він протікає в матриксі мітохондрій і складається з восьми послідовних реакцій.
Перша реакція каталізується цитрат-синтетазою, при цьому ацетил-КоА конденсується із щавелевооцтовою кислотою (оксалоацетатом), у результаті чого утворюється лимонна кислота (цитрат).

У другій реакції лимонна кислота піддається дегідратуванню з утворенням цисаконітової кислоти, що, приєднуючи молекулу води, переходить в ізолимонну кислоту (ізоцитрат).

Третя реакція - ізолимонна кислота окиснюється шляхом відщіплення двох атомів водню й перетворюється в щавелевобурштинову кислоту, що декарбоксилюється до α-кетоглутарової кислоти (α-кетаглутарат).

Четверта реакція: α-кетоглутарова кислота піддається окисному декарбоксилюванню під впливом складних ферментів До складу цих ферментів входять коферменти: (ТПФ, HSKoА, ліпоєва кислота, НАД, ФАД). Для реакції необхідні також іони Mg2+. У результаті окисного декарбоксилювання α-кетоглутарової кислоти утворюється сукциніл-КоА, що має макроергічний карбоксилтіоловий зв'язок, у якому резервується енергія окисного декарбоксилювання. При цьому виділяється друга молекула СО2 і НАДН2.

У ході п'ятої реакції сукциніл-КоА взаємодіє з неорганічним фосфатом, потім реагує з гуанідилдифосфатом (ГДФ) і перетворюється в бурштинову кислоту, а ГДФ, приєднуючи фосфат, переходить у ГТФ.

У результаті шостої реакції бурштинова кислота при участі ФАД окисляється у фумарову кислоту (фумарат). Остання гідратується з утворенням яблучної кислоти (сьома реакція). Під впливом НАД вона окисляється до щавлевооцтової кислоти (оксалоацетат) - восьма реакція, тобто до того субстрату, з якого починається цикл трикарбонових кислот.

Процес аеробного окиснювання вуглеводів відбувається з виділенням енергії (за рахунок НАДН2 і ФАДН,). Одна молекула НАДН2 виділяється на стадії декарбоксилювання пірувата з утворенням ацетил-КоА.
Окиснювання ізолимонної кислоти в щавелевобурштинову також супроводжується виділенням молекули НАДН2. При декарбоксилюванні α-кетоглутарової кислоти утворюється сукциніл-КоА й НАДН2. Сукциніл-КоА перетворюється в бурштинову кислоту з утворенням 1 молекули АТФ (за рахунок ГТФ). Дегідрування бурштинової кислоти відбувається з виділенням ФАДН2, а дегідрування яблучної - з виділенням НАДН2. Відомо, що 1 молекула НАДН2 еквівалентна 3 молекулам АТФ, а 1 молекула ФАДН2 - 2 молекулам АТФ.
Отже, з однієї тріози в аеробному циклі утворюється 4 молекули НАДН2 (12 молекул АТФ), 1 молекула ФАДН2 (2 молекули АТФ) і 1 молекула АТФ (ГТФ). Оскільки з однієї молекули глюкози утворюється 2 тріози, вихід енергії буде становити 15 х 2 = 30 молекул АТФ, а повне окиснювання 1 молекули глюкози в анаеробному і аеробному циклах складе 8 + 30 = 38 молекул АТФ.
Щавелевооцтова кислота, що утворилася, може вступати в реакцію з іншою молекулою ацетил-КоА й процес починається спочатку. Таким чином, певна кількість щавелевооцтової кислоти неодноразово бере участь у реакції й забезпечує окиснювання до СО2 і Н2О великої кількості оцтової кислоти, що утворюється з вуглеводів, ліпідів, білків й інших сполук. Але цей процес не нескінченний. Щавелевооцтова кислота в мітохондріях декарбоксилюється з утворенням піровиноградної кислоти, що у свою чергу перетворюється в ацетил-КоА.
Виділенням енергії не вичерпується значення циклу трикарбонових кислот. Роль його полягає також в тому, що при аеробному окиснюванні вуглеводів утворюються проміжні речовини, які використовуються для різних процесів біосинтезу. Так, α-кетоглутарова й щавелевооцтова кислоти можуть перетворюватися в амінокислоти (глутамінову й аспарагінову), необхідні для синтезу білків, азотистих основ, гормонів, нуклеотидів.
Цикл Кребса поставляє відбудовні еквіваленти в ланцюг дихальних ферментів, де потік електронів і протонів сполучений з утворенням АТФ. Цикл не може протікати швидше, ніж це дозволяє використання АТФ. Він регулюється за типом зворотного зв'язку: інгібується АТФ й активується АДФ, що накопичується в міру утилізації АТФ.
Пентозний цикл окиснювання вуглеводів
Подання про існування в тканинах тварин, рослин і мікробів іншого механізму окиснювання вуглеводів, що відрізняється від розглянутого вище, були розвинені на основі робіт головним чином Варбурга, Дікенса й В.А. Енгельгардта, що показали можливість окиснювання гексозомонофосфата в клітинах тварин (наприклад, в еритроцитах) без попереднього розщеплення на дві фосфотріози. Розбіжність шляхів окиснювання вуглеводів - класичного - лимоннокислого циклу Кребса й "пентозного" - починається на стадії утворення гексозомонофосфату. Якщо гексозомонофосфат (фруктозо-6-монофосфорний ефір) піддається ще раз фосфорилюванню й перетворюється у фруктозодифосфат, то в цьому випадку подальший розпад вуглеводів відбувається за звичайним гліколітичним шляхом з утворенням фосфотріоз і піровиноградної кислоти, яка "згоряє" потім у лимоннокислому "казані".
Якщо ж приєднання другої частки фосфату до гексозо-6-монофосфорному ефіру не відбувається, то глюкозомонофосфат може піддаватися прямому окислюванню з відщіпленням СО2 й утворенням пентозофосфату. Звідси відбулася й назва - пряме окиснювання вуглеводів у пентозному циклі.
Глюкозо-6-фосфат під впливом дегідрогенази піддається дегідруванню з утворенням спочатку 6-фосфоглюконолактону, а потім 6-фосфоглюконової кислоти.

6-фосфоглюконова кислота під впливом дегідрогенази, коферментом якої також є НАДФ, піддається дегідруванню й потім декарбоксилюванню з утворенням пентозофосфату: рибулозо-5-фосфату й однієї молекули СО2.
Реакція протікає в такий спосіб:

У ході зазначених перетворень 2 молекули НАДФ відновлюються в НАДФН2, які можуть бути використані для різних відбудовних синтезів.
Рибулозо-5-фосфат під впливом ізомерази легко перетворюється в біологічно важливий цукор - рибозо-5-фосфат. Між обома формами пентозофосфатів установлюється стан рухливої рівноваги:
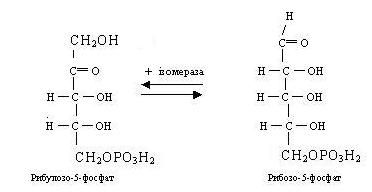
З рибулозо-5-фосфату під впливом ферменту епімерази може утворитися ще одна пентоза - ксилулозо-5-фосфат.

Гептоза - седогептулозо-7-фосфат - утворюється в пентозному циклі із ксилулозо-5-фосфата і рибозо-5-фосфату в результаті реакції, яка каталізується ферментом транскетолазою.

Тетроза- еритрозо-4-фосфат виникає в присутності трансальдолази в результаті розриву СѕС зв'язку в седогептулозі з перекиданням групи СН2ОН¾ СО¾ СНОН - на фосфогліцероловий альдегід:

Основний шлях окиснювання вуглеводів в організмі тварин здійснюється за звичайним гліколітичним шляхом (за схемою Ембдена-Мейергофа-Парнаса-Кребса). Тільки в окремих органах і тканинах, наприклад у жировій тканині, печінці, еритроцитах, надниркових залозах, лімфатичних вузлах і деяких інших тканинах має істотне значення окиснювання гексозофосфатів у пентозному циклі.
Види бродіння
У результаті життєдіяльності мікроорганізмів, що беруть участь у процесах обміну речовин, відбуваються різноманітні хімічні реакції, у результаті яких утворюються цінні речовини: спирти, кислоти, ефіри, вітаміни та інші. Вони широко використовуються в різних галузях промисловості, медицині, побуту. Багато біохімічних процесів, викликаних мікроорганізмами, знаходять застосування в харчовій і легкій промисловості.
Вивчення біохімічної діяльності мікроорганізмів дозволяє більш ефективно їх використати, правильно організувати технологічний процес. Разом з тим, виявлення сутності тих змін, які відбуваються в харчовій сировині й продуктах її переробки при спонтанному (мимовільному) розвитку в них мікроорганізмів, сприяє успішній боротьбі зі збудниками псування продуктів.
До анаеробних процесів, що протікають за участю мікроорганізмів, відносять: спиртове, молочнокисле, пропіоновокисле, маслянокисле, ацетонобутилове бродіння.
Спиртове бродіння може бути виражено сумарною реакцією:
С6Н12О6 + 2АДФ + 2ФН ↔ 2СН3 - СН2ОН + 2СО2 + 2АТФ + 2Н2О
На основі спиртового бродіння печуть хліб, виготовляють пиво, вино, спирт. Поряд з молочнокислим бродінням воно застосовується в технології одержання деяких молочнокислих напоїв (кумису, кефіру), а також при квашенні овочів. Мимовільно виникаюче спиртове бродіння у фруктових соках, сиропах, компотах, варенні викликає їхнє псування - заброджування. До збудників спиртового бродіння відносяться дріжджі (винні, пивні, пекарські, верхівкового і низового бродіння). Ферментами дріжджів зброджуються D-глюкоза, D-фруктоза, D-маноза, D-галактоза. Основними продуктами спиртового бродіння є етанол і СО2, побічними - гліцерол, оцтова й бурштинова кислоти, сивушні масла - суміші вищих спиртів (ізоамілового, ізобутилового, амілового, пропілового й ін.).
До стадії утворення пірувату (піровиноградної кислоти) гліколіз і спиртове бродіння протікають однаково. Потім при цьому виді бродіння піровиноградна кислота взаємодіє з тіамінпірофосфатом у присутності Mg2+, що приводить до її розщеплення на ацетальдегід і СО2.

На наступному етапі ацетальдегід відновлюється до етанолу алкоголь-дегідрогеназою (алкоголь НАД-оксидоредуктазой). Коферментом цього ферменту є НАДН2, що виділяється при дегідруванні проміжного фермент-субстратного комплексу, який з'являється при анаеробному окиснюванні глюкози. Спрямованість реакції залежить від рН середовища.

Утворення вищих спиртів пов'язане з азотним і вуглеводним обміном дріжджових клітин. Вищі спирти синтезуються при дезамінуванні й декарбоксилюванні амінокислот, у реакціях переамінування амінокислот з піровиноградною кислотою, у процесах синтезу амінокислот. Ці спирти беруть участь у формуванні аромату і смаку напоїв.
Побічний продукт реакції - гліцерол може перетворюватися в акролеїн, що заважає чистому процесу спиртового бродіння.
Молочнокисле бродіння - це перетворення цукру під дією ферментів молочнокислих бактерій у молочну кислоту.
Розрізняють гомо- і гетероферментативне молочнокисле бродіння. При гомоферментативному бродінні мікроорганізми зброджують гексози, в основному до молочної кислоти й лише в слідових кількостях утворюються побічні продукти: летучі кислоти, етанол, фумарова кислота й СО2.
Сумарне рівняння цього виду бродіння:
С6Н12О6 + 2АДФ + 2ФН ® 2СН3 - СНОН - СООН + 2АТФ.
При гетероферментативному молочнокислому бродінні поряд з молочною кислотою утворюються етанол, оцтова кислота і оксид карбогену.
Залежно від бактерій, що викликають даний процес, розрізняють два види бродіння: бродіння, що відбувається без виділення СО2, і бродіння, що супроводжується газовиділенням.
Молочнокисле бродіння використовують у виробництві різних молочних продуктів, при випічці хліба (особливо житнього), квашенні капусти і засолюванні огірків, силосуванні кормів, одержанні молочної кислоти.
Спонтанно виникаюче молочнокисле бродіння в молоці, вині, пиві, безалкогольних напоях приводить до їхнього псування (прокисанню, помутнінню, ослизненню та ін.).
Маслянокисле бродіння викликають анаеробні спороутворюючі бактерії. У харчовій промисловості воно, як правило, відіграє негативну роль. Так, масляна кислота, що утворюється при цьому бродінні, може зіпсувати молоко й молочні продукти. У спиртовому виробництві, що працює на зерновій або іншій крохмальвмісній сировині, при проростанні спор цих бактерій виділяється гідроген, що створює небезпеку вибуху. Крім того, масляна кислота гальмує ріст дріжджів, а отже порушує нормальний хід процесу. У спиртовому виробництві, де як сировина використовуються плоди і ягоди, "затори" можуть бути зіпсовані запахом масляної кислоти. Маслянокислі бактерії можуть викликати масову загибель картоплі й овочів, спучування сирів, псування консервів, прогіркання молока, вологого борошна, квашених овочів.
Рівняння маслянокислого бродіння:

Ефіри масляної кислоти відрізняються приємним ароматом. Їх використовують як ароматичні речовини в кондитерській і парфумерній промисловості і при виготовленні фруктових напоїв.
Пропіоновокисле бродіння - це перетворення цукру або молочної кислоти і її солей при участі пропіоновокислих бактерій у пропіонову й оцтову кислоти з виділенням СО2 і Н2О.
Сумарне рівняння реакції:

Цей вид бродіння є одним з найважливіших процесів при дозріванні сирів. Виділення вуглекислоти, що утворюється при цьому, викликає поява сирних "вічок" - порожнеч. Присутність пропіонової і оцтової кислот у сирі обумовлює його своєрідний гострий смак і запах.
Змішане кислотне бродіння відбувається за участю декількох видів мікроорганізмів (молочнокислі бактерії, оцтовокислі бактерії й ін.), при цьому із глюкози утворюються кислоти (молочна, оцтова, мурашина, бурштинова), етанол, гліцерол, СО2, ацетон і його похідні. Даний вид бродіння негативно позначається на виробництві багатьох продуктів. Наприклад, летуча оцтова кислота, що є одним з основних його речовин, погіршує якість настойок.
КОНТРОЛЬНІ ЗАВДАННЯ І ПИТАННЯ
1. Як відбуваються процеси переварювання вуглеводів у шлунково-кишковому тракті людини?
2. Як регулюється вміст цукру в крові людини?
3. Охарактеризуйте процеси біосинтезу глікогену.
4. У чому полягає суть гліколізу (анаеробне окиснювання глюкози)?
5. Напишіть реакції гліколізу.
6. Напишіть реакцію окисного декарбоксілювання піровиноградної кислоти.
7. Напишіть реакції циклу Кребса.
8. Який енергетичний вихід циклу Кребса?
9. Як протікає пентозофосфатний шлях окиснювання вуглеводів?
10. Які види бродіння лежать в основі одержання молочно-кислих продуктів, квашення овочів, виробництва хліба й хлібобулочних виробів, вин, чаю? Опишіть хімічні реакції різних видів бродіння.
Тести для контролю знань і самопідготовки
1. Як діє адреналін і симпато-адреналова система на рівень цукру в крові?
| o | а) підвищує (адреналова гіперглікемія), стимулюючи розпад глікогену |
| o | б) підвищує, гальмуючи синтез глікогену |
| o | в) не впливає |
| o | г) знижує рівень цукру в крові, гальмуючи синтез глікогену |
2. Як впливає інсулін на рівень цукру в крові?
| o | а) стимулює розпад глікогену, викликаючи гіперглікемію |
| o | б) стимулює синтез глікогену, утримуючи концентрацію цукру крові у фізіологічних межах |
| o | в) не впливає на синтез глікогену |
| o | г) стимулює синтез глікогену, викликаючи гіпоглікемію |
3. Як діє глюкагон на вуглеводний обмін?
| o | а) стимулює синтез глікогену, сприяючи виникненню гіперглікемії |
| o | б) стимулює розпад глікогену (антагоніст інсуліну) |
| o | в) не впливає на вуглеводний обмін |
| o | г) регулює гліконеогенез (синтез глікогену) |
4. Як впливають глікокортикоїди на обмін вуглеводів?
| o | а) стимулюють глікогенез із глюкози |
| o | б) стимулюють гліконеогенез |
| o | в) стимулюють гідроліз глікогену |
| o | г) стимулюють фосфороліз глікогену |
5. Чи впливають тиреоідні та статеві гормони на обмін вуглеводів?
| o | а) не впливають |
| o | б) стимулюють синтез глікогену |
| o | в) стимулюють енергетичний обмін глікогену |
| o | г) стимулюють глікогеноліз |
6. Яка роль синтезу глікогену?
| o | а) створення запасу вуглеводів |
| o | б) створення попередника гетерополісахаридів |
| o | в) глікоген- стартова сполука анаеробного і аеробного окислення вуглеводів |
| o | г) нагромадження запасу вуглеводів і стартової сполуки, яка легко гідролізується |
7. В яких тканинах в основному утворюється глікоген?
| o | а) мозку, серці, сполучній тканині |
| o | б) печінці, поперечно-смугастих м’язах |
| o | в) печінці, нирках, легенях, серці |
| o | г) у всіх без винятку тканинах |
8. Із яких моносахаридів синтезується глікоген?
| o | а) глюкоза |
| o | б) фруктоза |
| o | в) галактоза |
| o | г) пентоза |
9. Що таке гліконеогенез?
| o | а) синтез глікогену із глюкози |
| o | б) синтез глікогену із дисахаридів |
| o | в) синтез глікогену із проміжних продуктів метаболізму |
| o | г) синтез глікогену із ЖК |
10. Яким чином фруктоза бере участь в синтезі глікогену?
| o | а) участь не бере |
| o | б) спочатку перетворюється в глюкозу |
| o | в) безпосередньо включається в синтез глікогену |
| o | г) фосфорилюється АТФ і включається в синтез глікогену |
11. У чому полягає тлумачення терміну „гліколіз”?
| o | а) біологічне окислення глюкози в анаеробних умовах у тканинах тварин |
| o | б) аеробне окислення глюкози |
| o | в) спиртове бродіння та анаеробне окислення |
| o | г) ана- та аеробне окислення глюкози |
12. Яке сучасне визначення глікогенолізу?
| o | а) анаеробне окислення глюкози |
| o | б) аеробне окислення глюкози |
| o | в) анаеробне окислення глюкози та глікогену (глікогеноліз) у тканинах тварин |
| o | г) анаеробне окислення глікогену в тканинах тварин |
13. Який кінцевий продукт гліколізу?
| o | а) Н2О + СО2 |
| o | б) молочна кислота |
| o | в) піровиноградна кислота |
| o | г) спирт |
14. Як змінюється вміст цукру в крові під час збудження ЦНС?
| o | а) підвищується (психогенна гіперглікемія) |
| o | б) знижується (психогенна гіпоглікемія) |
| o | в) не змінюється |
| o | г) з’являється багато різних моносахаридів та інших похідних |
15. Як використовуються моносахариди в організмі людей?
| o | а) для синтезу глікогену і лактози |
| o | б) для синтезу гетерополісахаридів та інших похідних |
| o | в) для виробництва енергії в окисних процесах |
| o | г) забезпечують усі наведені функції |
16. Як перетворюється глікоген на першому етапі глікогенолізу?
| o | а) фосфоролізується за участю фосфорилази з утворенням глюкозо-6-фосфату |
| o | б) фосфоролізується з утворенням глюкозо-1-фосфату |
| o | в) гідролізується гамма-амілазою з утворенням вільної глюкози |
| o | г) гідролізується α-амілазою |
17. Яка сполука утворюється, коли в гліколіз безпосередньо включається глюкоза?
| o | а) глюкозо-1-фосфат (за участю глюкокінази) |
| o | б) глюкозо-6-фосфат (за участю глюкокінази) |
| o | в) глюкозо-1,6-дифосфат (за участю АТФ) |
| o | г) фруктозо-6-фосфат (за участю гексокінази) |
18. Як перетворюється фруктозо-6-фосфат під дією фосфофруктокінази?
| o | а) за рахунок АТФ фруктозо-6-фосфат за наявності іонів магнію перетворюється на фруктозо-1,6-дифосфат |
| o | б) за рахунок гідролізу утворюється вільна фруктоза і Фн. |
| o | в) фруктозо-6-фосфат за наявності іонів магнію перетворюється на глюкозо-6-фосфат |
| o | г) фруктозо-6-фосфат перетворюється на галактозо-6-фосфат |
19. На які сполуки перетворюється фруктозо-1,6-дифосфат під впливом альдолази?
| o | а) на фруктозо-1-фосфат |
| o | б) на фруктозо-2,6-фосфат |
| o | в)на вільну фруктозу |
| o | г) на діоксиацетонфосфат і гліцеринальдегідфосфат |
20. Яких подальших перетворень зазнають тріозофосфати у ході гліколізу?
| o | а) діоксиацетонфосфат за участю тріозофосфатизомерази перетворюється на гліцеринальдегідфосфат |
| o | б) гліцеринальдегідфосфат ізомеризується в діоксиацетонфосфат |
| o | в) два тріозофосфати конденсуються у глюкозу |
| o | г) два тріозофосфати конденсуються у фруктозо-1,6-дифосфат |
21. Яка сполука є кінцевим продуктом гліколізу?
| o | а) гліцериновий альдегід |
| o | б) гліцеринова кислота |
| o | в) СО2 і Н2О |
| o | г) молочна кислота |
22. Яка макроергічна сполука нагромаджується під час перетворення фосфоенолпірувату?
| o | а) АТФ |
| o | б) креатинфосфат |
| o | в) піровиноградна кислота |
| o | г) гліцерат-2-фосфат |
23. Яка сполука утворюється внаслідок окисного декарбоксилування пірувату?
| o | а) лактат |
| o | б) гліцеральфосфат |
| o | в) ацетил-КоА |
| o | г) СО2 і Н2О |
24. Де нагромаджується Н+ анаеробного дегідрування?
| o | а) на ФАД.Н2 |
| o | б) на НАДН.Н+ |
| o | в) на проміжних метаболітах |
| o | г) на цитохромах |
25. Скільки молекул НАДН2 утворилося внаслідок окислення молекули глюкози до гліцеро-1,3-дифосфату
| o | а) одна |
| o | б) дві |
| o | в) три |
| o | г) чотири |
26. Який метаболіт є кінцевим акцептором 2Н під час гліколізу?
| o | а) гліцеринальдегід-3-фосфат |
| o | б) молочна кислота |
| o | в) піровиноградна кислота |
| o | г) оксиген |
27. Що таке аеробне окислення?
| o | а) окислення за наявності кисню |
| o | б) окислення в безкисневому середовищі |
| o | в) окислення киснем |
| o | г) окислення шляхом дегідрування |
28. Які речовини окислюються в аеробному процесі (диханні) за участю вуглеводів?
| o | а) піровиноградна кислота, янтарна та інші органічні кислоти |
| o | б) глікоген, глюкоза, молочна кислота |
| o | в) молочна та оцтова кислоти |
| o | г) піровиноградна кислота |
29. Якими реакціями починається аеробне окислення вуглеводів?
| o | а) окисленням молочної кислоти |
| o | б) перетворенням піровиноградної кислоти |
| o | в) перетворенням глікогену і глюкози до піровиноградної кислоти, як і під час гліколізу |
| o | г) окисленням етилового спирту |
30. На якому етапі з’являються розбіжності процесів аеробного та анаеробного окислення вуглеводів?
| o | а) на етапі подальшого перетворення піровіноградної кислоти. Під час анаеробного окислення вона відтворюється на молочну кислоту, аеробно – декарбоксилюється з утворенням ацетил-КоА |
| o | б) гліколіз починається з окислення глюкози, адсорбне окислення- глікогену |
| o | в) на етапі окислення фосфорилювання гліцеральфосфату |
| o | г) на етапі термінального окислення НАДН2 |
31. На якому етапі клітини нагромаджують більшість АТФ в аеробних умовах?
| o | а) на етапі окислення глюкози до піровиноградної кислоти |
| o | б) на етапі окислення гліцервль-3-фосфату |
| o | в) за рахунок окислення молочної кислоти до пірувату |
| o | г) за рахунок окислення пірувату в ЦТК (аеробний цикл) |
32. Які вуглеводи утворюються в тканинах людини?
| o | а) крохмаль, гліцерин, діоксиацетон |
| o | б) глікоген, глюкоза, гліцериновий альдегід, діоксиацетон, Д-рибоза |
| o | в) глікоген, амілоза, амілопектин і хітин |
| o | г) целюлоза, крохмаль, маноза, галактоза |
33. Яка сполука забезпечує перебіг глікогенезу і яким ферментом каталізується цей процес?
| o | а) глюкозо-1-фосфат, перенесений на УДФ-глюкозу за наявності затравочної кількості глікогену і глікогенсінтетази |
| o | б) глюкозо-6-фосфат за участю фруктозо-1,6-дифосфату |
| o | в) фруктозо-6-фосфат за наявності затравочної кількості глікогену і синтетаз |
| o | г) лимонна та ізолимонна кислоти |
34. Що таке гліконеогенез?
| o | а) розщеплення глікогену в печінці та м’язах |
| o | б) анаеробне окислення глікогену |
| o | в) аеробне окислення глікогену |
| o | г) синтез глікогену із продуктів обміну |
35. На яку сполуку перетворюється глюкозо-6-фосфат у гліколізі?
| o | а) на фруктозо-6-фосфат |
| o | б) на глюкозо-1-фосфат |
| o | в) на глюкозо-1,6-дифосфат |
| o | г) на тріозофосфат |
36. Куди переноситься глюкоза з уридилдифосфатглюкози під дією глікогенсинтетази?
| o | а) на редукуючий кінець амілозного ланцюга глікогену |
| o | б) на нередукуючий кінець амілозного ланцюга |
| o | в) на амілозний ланцюг з утворенням а(6-1) зв’язків |
| o | г) на ділянку утворення а(4-1) і а(6-1) зв’язків |
37. Яким чином утворилася така сполука?
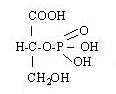
| o | а) ізомеризацією гліцеро-3-фосфату в гліцеро-2-фосфат |
| o | б) окисним фосфорилюванням |
| o | в) фосфорилюванням |
| o | г) етерифікацією Фн |
38. Яка сполука має таку формулу?

| o | а) 3 фосфогліцерінова кислота |
| o | б) гліцеро-1- фосфат |
| o | в) гліцеро-2-фосфат |
| o | г) диоксиацетон фосфат |
39. Які сполуки мають такі формули:

| o | а) фосфоенолпіровиноградна, молочна, піровиноградна кислоти |
| o | б) піровиноградна, фосфоенолпіровиноградна, молочна кислота |
| o | в) молочна, фосфоенолпіровиноградна, піровиноградна кислота |
| o | г) фосфоенолпіровиноградна, піровиноградна, молочна кислота |
40. Що таке обмін вуглеводів?
| o | а) перетравлення, всмоктування вуглеводів, біосинтез із АК, гліцерину і молочної кислоти |
| o | б) перетравлення, всмоктування і виділення продуктів їх обміну - H2О і CO2 |
| o | в) перетравлення вуглеводів до моносахаридів, всмоктування їх в кров, взаємоперетворення, анаеробне і аеробне окислення і синтез вуглеводів у тканинах |
| o | г) розщеплення харчових вуглеводів у шлунково-кишковому тракті, гліколіз і синтез за рахунок гліконеогенезу |
41. Де і якими ферментами перетравлюється крохмаль (глікоген)?
| o | а) переважно у ротовій порожнині, тонкому кишечнику α-амілазою до декстринів і мальтози |
| o | б) у шлунку пепсином |
| o | в) у кишечнику трипсином і хімотрипсином |
| o | г) у кишечнику ліпазою |
РОЗДІЛ 12. ОБМІН ЛІПІДІВ
12.1. РОЗЩЕПЛЕННЯ ЛІПІДІВ У ТРАВНОМУ ТРАКТІ ЛЮДИНИ
Розщеплення ліпідів у травному тракті людини має кілька стадій. Для цього процесу необхідні ліполітичні ферменти (і відповідні умови для їхньої діяльності) і емульгатори (детергенти). За сучасною класифікацією ліполітичні ферменти відносяться до групи гідролаз, які каталізують розщеплення різних ліпідів. У травному тракті людини субстратами гідролітичного розщеплення є триацилгліцероли, фосфоліпіди, ефіри холестеролу.
Оптимальна умова для дії ліполітичних ферментів - рН 7,8-8,2. Гідролітичному розщепленню в шлунку піддаються тільки емульговані жири. Такі ліпіди містяться в молоці і молочних продуктах, яєчному жовтку, майонезах. Всі інші жири їжі мають потребу в емульгаторах, що знижують поверхневий натяг і перешкоджають склеюванню жирових крапель.
Емульгатори мають гідрофільні і гідрофобні групи, вони оточують кожну краплю жиру таким чином, що гідрофільні групи звертаються до води, а гідрофобні - до жиру. Основними емульгаторами жирів у травному тракті людини є солі жовчних кислот.
У порожнині рота переварювання ліпідів не відбувається через відсутність ліпаз. У шлунку йде незначний гідроліз емульгованих жирів під дією малоактивної ліпази шлункового соку. Основна кількість харчових жирів гідролізується в тонкому кишечнику під дією ліпази, що утворюється в підшлунковій залозі (це можливо тільки після емульгування жирів).
Емульгування жирів відбувається в порожнині кишечнику під впливом дрібних пухирців вуглекислого газу, що рясно виділяються при нейтралізації соляної кислоти харчової кашки бікарбонатами підшлункового і кишкового соків. У процесі перистальтики кишечнику жири роздрібнюються на дуже дрібні краплі, що емульгуються при участі парних жовчних кислот і моноацилгліцеролів. Основну роль при цьому грають солі жовчних кислот (мила), що виділяються з жовчю в просвіт кишечнику. Вони адсорбуються на поверхні крапель жиру, утворюють на них найтоншу плівку, що перешкоджає злиттю крапельок у більш великі краплі. Разом з тим жовчні кислоти різко зменшують натяг на поверхні двох фаз - води і жиру, що сприяє дробленню його крапель на більш дрібні. При цьому утвориться тонка емульсія (діаметр часток не перевищує 0,5 мкм), що полегшує ферментативний гідроліз жиру. Одночасно жовчні кислоти активують ліпазу.
Жовчні кислоти являють собою похідні холанової кислоти, що синтезується з холестеролу. У жовчі людини містяться в основному похідні холанової кислоти: холева, дезоксихолева, глікохолева, хенодезоксихолева, таурохолєва і літохолева кислоти, причому головним чином їх натрієві солі, що складаються з холевої і дезоксихолевої кислот, глікоколу і таурину.

Велика частина емульгованого жиру піддається гідролітичному розщепленню під дією ліпаз з утворенням гліцеролу і вищих жирних кислот.
Ліпази бувають двох типів: одна розщеплює ефірні зв'язки триацилгліцеролів у положеннях 1 і 3, інша - у положенні 2. У розщепленні моноацилгліцеролів бере участь також ліпаза, що міститься в кишковому соку.
Гідроліз є першою фазою обміну жирів. Він протікає східчасто, причому спочатку розщеплюються зовнішні складноефірні зв'язки. a-моноацилгліцероли, що утворилися, всмоктуються стінкою кишечнику. Потім вони або йдуть на ресинтез триацилгліцеролів у кишковій стінці, або розпадаються під дією неспецифічних естераз. Жирні кислоти, що виділилися з розщеплених гліцеролів, погано розчиняються у воді і всмоктуються ворсинками кишечнику лише після взаємодії з жовчними кислотами з утворенням парних розчинних комплексів. В епітеліальних клітинах ворсинок кишечнику відбувається їхнє розщеплення на жовчні і жирні кислоти. Жовчні кислоти знову безпосередньо надходять у просвіт кишечнику або проходять більш складний шлях: кров - печінка - жовчний міхур – жовч.
Постійна циркуляція жовчних кислот забезпечує велику кількість всмоктуваних жирів при порівняно обмеженому виробленні печінкою жовчних кислот (2,8-3,5 г у добу).
Переварювання ліпідів відбувається як у порожнині кишок (порожнинне травлення), так і на слизовій оболонці тонкої кишки (пристінкове або контактне травлення). Зокрема, на поверхні клітин адсорбується ліпаза соку підшлункової залози (панкреатична), яка каталізує гідроліз жирів.
З епітеліальних клітин стінки кишечнику жири у виді дрібних жирових крапель, оточених білками (хіломікрони), попадають у лімфу.
Хіломікрони відносяться до складних ліпідів. За будовою це сферичні частки (100-5000 нм), зовнішня оболонка яких утворена білками, а внутрішня - ліпідами і їхніми похідними. Хіломікрони - найбільш великі з ліпопротеїнів. До їхнього складу входить 98-99 % ліпідів і 1- 2 % білка. Утворюючись в клітинах слизової оболонки кишечнику, вони забезпечують транспорт ліпідів (триацилгліцеролів) з кишечнику в лімфу.
Хіломікрони через грудну лімфатичну протоку надходять у кровоток і транспортуються в "жирові депо" і печінку.
Таким чином, за допомогою хіломікронів здійснюється транспорт із кишечнику в кров екзогенних триацилгліцеролів, холестеролу і частково фосфоліпідів. Через 1-2 год після прийому їжі спостерігається аліментарна гіперліпемія, максимальної величини вона досягає через 4-6 годин після прийому жирної їжі. Через 10-12 годин вміст триацилгліцеролів повертається до норми, а хіломікрони зникають із кров'яного русла, тому що розпадаються на більш дрібні частки під впливом ліпопротеїнліпази (фактор просвітління) внутрішньої оболонки стінок судин.
Ліпопротеїнліпаза не тільки розщеплює хіломікрони, але і гідролізує триацилгліцероли, що виділилися при цьому, на гліцерол і жирні кислоти, що можуть безпосередньо надходити в клітки і піддаватися процесам розпаду. Частина жирних кислот попадає в кров, де адсорбується альбуміном сироватки, потім переноситься до органів і включається в процеси обміну (рис. 5.1).
Частина жирів безпосередньо всмоктується в кров, минаючи лімфатичну систему, і надходить у печінку. Через деякий час з печінки жири переходять у периферичне депо: підшкірну клітковину, сальник, брижі. Жири з "жирових депо" осідають в інших тканинах, головним чином у печінці, де піддаються окисному розщепленню до кінцевих продуктів обміну речовин. Транспорт жирів у крові забезпечують білки плазми: b- і g-глобуліни, альбуміни.

Рис 5.1. Модель ліпопротеїнів
1 - ефіри холестерола; 2 - триацилгліцероли; 3 - фосфоліпіди; 4 - холестерол; 5- полярний шар
Холестерол попадає в шлунково-кишковий тракт людини переважно з яєчним жовтком, м'ясом, печінкою, мізками. З їжею людина одержує щодня 0,1-0,3 г холестеролу у вільному виді або у виді його ефірів. Останні при участі ферменту панкреатичного соку - холестеролестерази розщеплюються на холестерол, що може всмоктуватися у виді комплексу з жовчними кислотами, і жирні кислоти.
Фосфоліпіди, зокрема лецитини, під впливом відповідних гідролаз, розщеплюються на гліцерол, вищі жирні кислоти, холін і фосфорну кислоту. Компоненти фосфоліпідів усмоктуються кишковою стінкою і надходять у кров (фосфорна кислота в основному у виді натрієвих і калієвих солей).
З продуктів гідролізу харчових ліпідів у клітинах кишкового епітелію ресинтезуються ліпіди, специфічні для певного виду тварини.
Важлива роль в обміні жирів належить печінки. Її ферментативні системи каталізують переважну більшість реакцій метаболізму ліпідів. У печінці синтезуються триацилгліцероли, що або затримуються в ній, або у виді ліпопротеїнів надходять у кров.
Розрізняють два види ліпопротеїнів, що утворюються в печінці: пре-b-ліпопротеїни (ліпопротеїни дуже низької щільності - ЛПОНП) і a-ліпопротеїни (ліпопротеїни високої щільності - ЛПВП). У плазмі крові людини з пре-b-ліпопротеїнів утворюються b - ліпопротеїни (ліпопротеїни низкої щільності - ЛПНП).
З усіх ліпопротеїнів плазми крові атерогеністю володіють b -ліпопротеїни низької і дуже низької щільності. Вони здатні проникати всередину судинної стінки з плазми крові, будучи первинним субстратом, що викликає атеросклеротичну поразку артерій. При атеросклерозі рівень холестеролу в крові підвищується в 2-5 разів (іноді до 13 ммоль/л). Причиною гіперхолестеролемії є порушення рівноваги між кількістю холестеролу, що розпадається, і синтезованого холестеролу. Основну роль у виникненні гіперхолестеролемії належить ендогенному холестеролу. Збільшення цього показника відзначено при надлишковому споживанні жирів і вуглеводів і порушенні їх утилізації.
Холестерол з ацетил-КоА також синтезується в печінці. Активність цього процесу залежить від кількості екзогенного холестеролу, що надходить з їжею. Біосинтез холестеролу в печінці регулюється за принципом негативного зворотного зв'язку: чим більше його надходить з їжею, тим менше синтезується в печінці, і навпаки.
Частина синтезованого холестеролу перетворюється в жовчні кислоти і виділяється з організму з жовчю, крім того він використовується для синтезу стероїдних гормонів і інших сполук. У печінці відбувається його взаємодія з жирними кислотами з утворенням ефірів, що надходять у кров.
У печінці відбувається розпад і синтез фосфоліпідів. Для їхнього синтезу, крім гліцеролу і вищих жирних кислот, необхідні неорганічні фосфати і азотисті основи, зокрема холін. Неорганічні фосфати в печінці містяться в достатній кількості, а холіну мало, тому він обов'язково повинний надходити з їжею.
При дефіциті холіну синтез фосфоліпідів з нейтрального жиру різко знижується або зовсім припиняється, і нейтральний жир відкладається в печінці, тобто відбувається жирова інфільтрація цього органа, що грозить перейти в її жирову дистрофію. Запобігають цьому ліпотропні речовини (холін, метіонін і інші джерела метильних груп), що містяться в сирі й інших молочних продуктах, м'ясі та ін.
12.2. ОБМІН ЛІПІДІВ У ТКАНИНАХ
Обмін ліпідів у тканинах є біологічно найбільш важливим етапом їхнього перетворення. На цій фазі відбувається асиміляція ліпідів у виді пластичного матеріалу і розщеплення їх з вивільненням енергії.
Головним ендогенним джерелом ліпідів, що грають роль метаболічного палива, служить резервний жир, що міститься в протоплазмі клітин у виді крапельок. Для цієї мети використовуються також фосфоліпіди мембран.
У "жирових депо" при участі тканинних ліпаз відбувається гідроліз простих жирів на глицерол і вільні жирні кислоти. Гліцерол фосфорилюється за рахунок АТФ, через ряд проміжних реакцій перетворюється у фосфогліцероловий альдегід, що потім окисляється в процесі гліколізу до фосфогліцеролової і піровиноградної кислот. Остання, піддаючи окисному декарбоксилюванню, перетворюється в ацетил-КоА, що у циклі трикарбонових кислот окисляється до СО2 і Н2О.
У виді комплексу з альбумінами вільні жирні кислоти зі струмом крові попадають в органи і тканини, де комплекс розпадається, а жирні кислоти або піддаються b-окислюванню, або використовуються в синтезі триацилгліцеролів, холестеролу, гліцерофосфоліпідів, сфінголіпідів і т.д.
Не нашли, что искали? Воспользуйтесь поиском: