ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Г — головка; Н — ножка
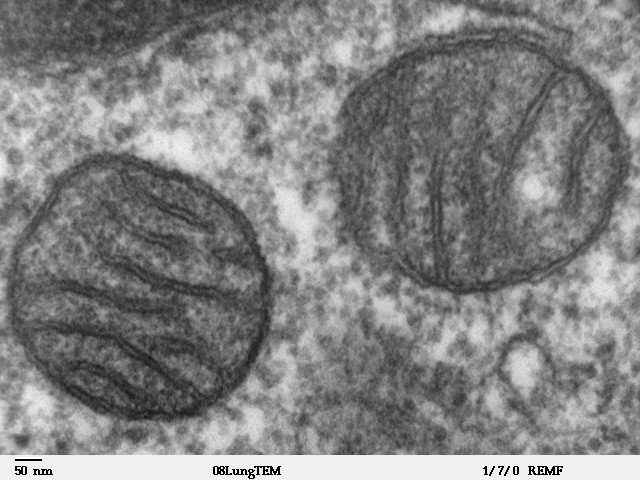
Митохондриальная ДНК образует собственный геном митохондрий, на который приходится около 1 % общего содержания ДНК в клетке. Наружная мембрана образована по принципу цитолеммы. Она обладает высокой проницаемостью и содержит молекулы транспортных белков, на ее поверхности содержатся рецепторы. Внутренняя мембрана обладает низкой проницаемостью для мелких ионов, содержит транспортные белки, ферменты дыхательной цепи и АТФ-синтетазные комплексы. Кристы внутренней мембраны митохондрий большинства клеток имеют пластинчатую форму и лишь у некоторых клеток содержатся везикулярные или трубчатые кристы (в эндокринных клетках, продуцирующих стероидные гормоны).
Митохондрии размножаются делением, живут около 10 дней, после чего подвергаются разрушению аутофагией. Они чувствительны к изменениям метаболизма клетки — трофики, рH среды, осмотического давления. Митохондрии первыми среди клеточных структур реагируют на воздействия повреждающих факторов.
Функции:
Энергетические станции клетки (синтез АТФ на пластинчатых кристах).
Дыхательный центр клетки.
Участие в биосинтезе стероидов (на везикулярных кристах, с участием кислорода).
Участие в окислении жирных кислот.
Участие в синтезе нуклеиновых кислот(собственных).
Жгутик


Поперечный срез жгутика эукариот в ундулаподии 1A и 1B — A и B микротрубочки периферического дублета, 2 — центральная пара микротрубочек и центральная капсула, 3 — динеиновые ручки, 4 — радиальная спица, 5 — нексиновый мостик, 6 — клеточная мембрана.
Жгу́тик
органоид движения, обеспечивающий перемещение клетки в жидкой среде или циркуляцию жидкой среды вокруг клетки. Имеется у жгутиконосцев, зооспор водорослей и грибов, сперматозоидов, у губок и др. Жгутик бактерий – изогнутая нить из 3—11 скрученных фибрилл из белка флагеллина, не способных изгибаться. Движутся, вращаясь в мембране. Жгутик эукариот – трубчатый вырост клетки, покрытый мембраной, содержит аксонему – пучок белковых фибрилл, состоящих из 2 центральных и 9 пар периферических микротрубочек. В основании такого жгутика лежат два взаимно перпендикулярных базальных тельца. Движутся волнообразно или воронкообразно, используя энергию АТФ.
. (Источник: «Биология. Современная иллюстрированная энциклопедия.» Гл. ред. А. П. Горкин; М.: Росмэн, 2006.)
Клеточный центр
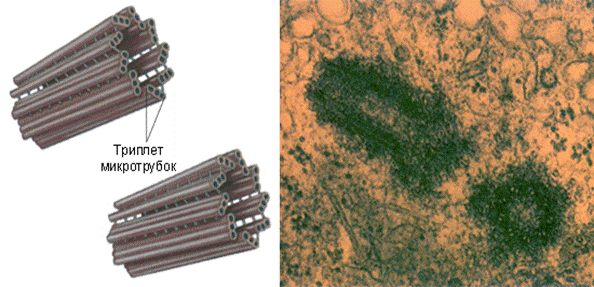
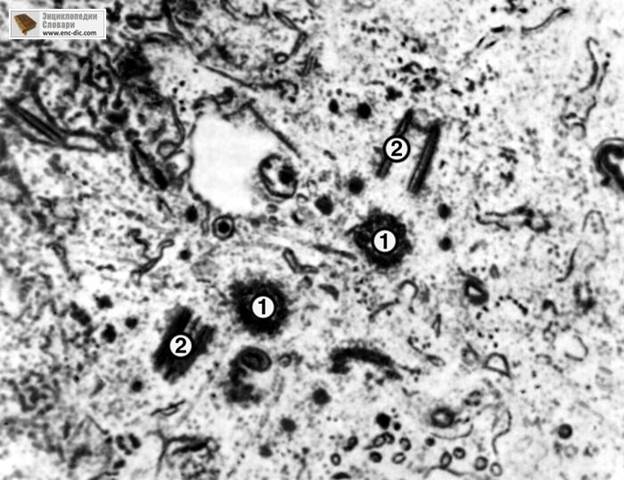 Рис.3
Рис.3
Электронограмма клеточного центра (две центриоли в
конце G1-периода клеточного цикла):
Рис. 5. Электронограмма клеточного центра (две центриоли в конце G1-периода
клеточного цикла):
1 — центриоли в поперечном сечении;
2 — центриоли в продольном сечении.
Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС) была открыта К.Р. Портером в 1945 г. Эта мембранная органелла общего назначения представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих как бы мембранную сеть внутри цитоплазмы. Различают два типа — гранулярную (шероховатую) и гладкую эндоплазматическую сеть.
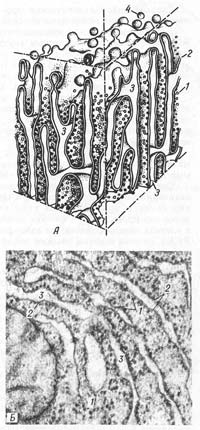 Рис. 8. Строение гранулярной эндоплазматической сети. А — схема; Б — электронная микрофотография участка среза печеночной клетки. 1 — рибосомы; 2 — пластинки; 3 — внутренние полости цистерн; 4 — отщепляющиеся мембранные пузырьки, лишенные рибосом.
Рис. 8. Строение гранулярной эндоплазматической сети. А — схема; Б — электронная микрофотография участка среза печеночной клетки. 1 — рибосомы; 2 — пластинки; 3 — внутренние полости цистерн; 4 — отщепляющиеся мембранные пузырьки, лишенные рибосом.
Гранулярная эндоплазматическая сеть (reticulum endoplasmaticum granulosum) представлена замкнутыми мембранами, которые образуют уплощенные мешки, цистерны, трубочки. Ширина полостей цистерн значительно варьирует в зависимости от функциональной активности клетки. Наименьшая ширина их — около 20 нм, но они могут достигать диаметра в несколько микрометров. Отличительной чертой мембран гранулярной ЭПС является то, что они со стороны гиалоплазмы покрыты рибосомами (рис. 8).
Гранулярная эндоплазматическая сеть бывает представлена редкими разрозненными цистернами или их локальными скоплениями. Первый тип гранулярной эндоплазматической сети, характерен для малоспециализированных ктеток или для клеток с низкой метаболической активностью. Скопления эндоплазматической сети являются принадлежностью клеток, активно синтезирующих секреторные белки. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазматическая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки.
Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков — ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они часто модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. В ряде случаев внутри самих канальцев или вакуолей гранулярной эндоплазматической сети может происходить модификация белков, например связывание их с сахарами (глюкозилирование), или конденсация синтезированных белков с образованием крупных агрегатов — секреторных гранул.
В гранулярной эндоплазматической сети происходит синтез мембранных интегральных белков (см. часть 2), которые встраиваются в толщу мембраны.
 Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50—100 нм.
Гладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети (при освобождении ее от рибосом).
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих такие категории липидов, как стероиды, например, в клетках коркового вещества надпочечников, в сустентоцитах семенников.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматическая сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при ряде отравлений в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь занятые гладким эндоплазматическим ретикулумом.
Не нашли, что искали? Воспользуйтесь поиском: