ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Биосинтез насыщенных жирных кислот
Синтез жиров осуществляется главным образом из углеводов, поступивших в избыточном количестве и не используемых для пополнения запаса гликогена. Кроме того, в синтезе участвуют и некоторые аминокислоты. Накоплению жиров способствует и избыток пищи.
Строительным блоком для синтеза жирных кислот в цитозоле клетки служит ацетил-КоА, который в основном поступает из митохондрий. Ацетил Ко-А самостоятельно не может диффундировать в цитозоль клетки, так митохондриальная мембрана непроницаема для него. Вначале внутримитохондриальный ацетил-КоА взаимодействует с оксалоацетатом, в результате чего образуется цитрат. Реакция катализируется ферментом цитрат-синтазой. Образовавшийся цитрат переносится через мембрану митохондрий в цитозоль при помощи специальной трикарбоксилаттранспортирующей системы.
В цитозоле цитрат реагирует с НS-КоА и АТФ, вновь распадаясь на ацетил-КоА и оксалоацетат. Эта реакция катализируется АТФ-цитрат-лиазой. Уже в цитозоле оксалоацетат при участии цитозольной малатдегидрогеназы восстанавливается до малата. Последний при помощи дикарбоксилаттранспортирующей системы возвращается в митохондриальный матрикс, где окисляется до оксалоацетата.

Имеются два типа синтазных комплексов, катализирующих биосинтез жирных кислот, оба находятся в растворимой части клетки. У бактерий, растений и низших форм животных, таких как эвглена, все индивидуальные ферменты синтазной системы находятся в виде автономных полипептидов; ацильные радикалы связаны с одним из них, получившим название «ацилпереносящий белок» (АПБ). У дрожжей, млекопитающих и птиц синтазная система представляет собой полиферментный комплекс, который нельзя разделить на компоненты, не нарушив его активности, а АПБ является частью этого комплекса. Как АПБ бактерий, так и АПБ полиферментного комплекса содержит пантотеновую кислоту в виде 4/-фосфопантетеина. В синтетазной системе АПБ выполняет роль СоА. Синтазный комплекс, катализирующий образование жирных кислот, является димером. У животных мономеры идентичны и образованы одной полипептидной цепью, включающей 6 ферментов, катализирующих биосинтез жирных кислот, и АПБ с реакционноспособной SH-группой, принадлежащей 4/-фосфопантетеину. В непосредственной близости от этой группы расположена другая сульфгидрильная группа, принадлежащая остатку цистеина, входящего в состав 3-кетоацил-ситазы (конденсирующего фермента), которая входит в состав другого мономера. Поскольку для проявления ситазной активности необходимо участие обеих сульфгидрильных групп, синтазный комплекс активен только в виде димера.
Первой реакций биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуется бикарбонат, АТФ, ионы марганца. Катализирует реакцию ацетил – КоА-карбоксилаза. Фермент относится к классу лигаз и содержит в качестве простетической группы биотин.
Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II-перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА:
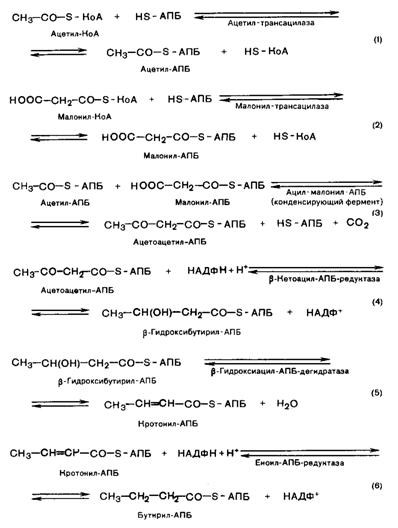
Малонил-КоА переходит в комплекс с SH-АПБ при участии фермента малонил-трансцилазы. В следующей реакции происходит взаимодействие ацетил-S-АПБ и малонил-S-АПБ. Происходит выделение карбоксильной группы малонил-S-АПБ в виде СО2. Ацетоацетил-S-АПБ при участии НАДФ+-зависимой редуктазы восстанавливается с образованием b-гидроксибутирил-S-АПБ. Далее реакция гидратации b-гидроксибутирил-S-АПБ приводит к образованию кротонил-b-гидроксибутирил-S-АПБ, который восстанавливается НАДФ+-зависимой редуктазой с образованием бутирил-S-АПБ. Далее рассмотренный цикл реакций повторяется: полученный бутирил-S-АПБ реагирует с другой молекулой малонил-S-АПБ с выделением молекулы СО2 (рис. 42).

Рис. 42. Биосинтез жирных кислот
В случае синтеза пальмитиновой кислоты (С16) необходимо повторение шести реакций, началом каждого из циклов будет присоединение молекулы малонил-S-АПБ к карбоксильному концу синтезируемой цепи жирной кислоты. Таким образом, присоединяя одну молекулу малонил-S-АПБ, углеродная цепь синтезируемой пальмитиновой кислоты увеличивается на два углеродных атома.
Синтез пальмитиновой кислоты завершается гидролитическим расщеплением SН-АПБ от пальмитил-S-АПБ при участии фермента деацилазы:
СН3-(СН2)14 –СО – S-АПБ + НО2 ® СН3-(СН2)14 –СООН + НS-АПБ
Синтез пальмитиновой кислоты является основным в синтезе жирных кислот организма. Свободная пальмитиновая кислота затем при участии тиокиназы превращается в пальмитил-КоА. Пальмитил-КоА может с участием карнитина проникать в митохондрии для синтеза жирных кислот с более длинной углеродной цепью.
Не нашли, что искали? Воспользуйтесь поиском: