ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Поведение реагента.
Обратившись теперь к поведению реагента, можно также обнаружить, что оно врожденное. Птенец серебристой чайки стремится клюнуть конец родительского клюва сразу же, без всякого научения. Выращенный в изоляции самец колюшки агрессивно реагирует на других самцов и ухаживает за самками, хотя у него не было никакой возможности этому научиться. Другими словами, речь идет не только о врожденной способности выполнять некоторые двигательные программы, но и о восприимчивости к особым запускающим и направляющим действия стимулам.
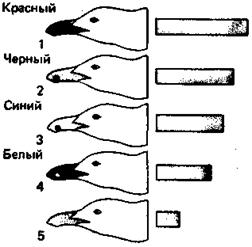
Рис. 48. Модели голов серебристой чайки с пятнами разного цвета на клюве (1—4) и без пятна (5). Полоски справа отражают частоту выпрашивающей реакции, вызванной соответствующей моделью.
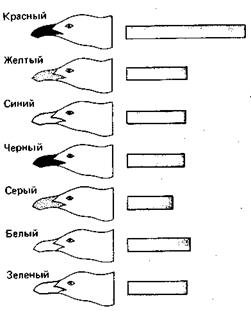
Рис. 49. Модели головы серебристой чайки с клювами разного цвета. Красный клюв вызывает больше реакций, чем любой другой, в том числе и желтый.
Способность реагировать на сигналы специально изучалась во многих случаях. Некоторые из результатов уже были описаны в предыдущих главах. Рассмотрим теперь более подробно выпрашивающее поведение птенца серебристой чайки, поскольку в данном случае точно известно, на какие стимулы он реагирует. Можно стимулировать выпрашивание, предъявляя ему плоские картонные макеты родительской головы (вкладка 6). На конце клюва у взрослой серебристой чайки находится красное пятно, четко выделяющееся на желтом окружающем фоне. Когда этого пятна на [80] макете нет, птенец реагирует гораздо слабее, чем в случае «нормального» макета с пятном (рис. 48). Если предъявлять их по очереди нескольким птенцам, среднее число реакций на макет без пятна оказывается в четыре раза меньше. Если это пятно не красное, стимулируется промежуточное число реакций. Оно зависит от степени контраста между цветом пятна и фона. Аналогичным образом, т. е. сравнивая реакции птенца на разные макеты, можно изучить влияние желтой фоновой окраски клюва. Удивительно, но от нее поведение птенцов почти не зависит, за исключением того, что на красный клюв они реагируют вдвое чаще, чем на любой другой (рис. 49). Клюв естественного желтого цвета вызывает не больше реакций, чем белый, черный, зеленый или синий. Окраска головы также не играет роли: можно было бы ожидать, что белая голова вызовет больше реакций, чем черная или зеленая, но это вовсе не так. Не существенна и форма головы; неважно даже, есть ли она вообще — был бы клюв. Однако птенцы прекрасно видят голову родителя, поскольку случайно могут клюнуть в основание клюва или даже в красное веко кормящей тики. Когда они голодны, для них существует только одно — родительский клюв с красным пятном. Кроме того, он должен быть тонким и длинным, направленным вниз, находиться как можно ближе к птенцу и пониже над землей. Вот и все стимулы; остальное для птенца не имеет значения. Поведение и окраска родителей прекрасно удовлетворяют этим требованиям, так сказать, полностью соответствуя ожиданиям потомства. Взрослая чайка подходит к птенцу, предъявляет ему свой клюв в почти вертикальном положении вниз концом, на котором находится красное пятно. Это совпадение признаков родителей и стимулов, на которые реагирует птенец, удивительно, если вспомнить, что он «не знает», как выглядит родитель или как он себя ведет.
У многих других изученных животных также наблюдалось, [81] что реагент, точно так же как и птенец чайки, отвечает всего на несколько стимулов, исходящих от агента. Уже говорилось, что агрессивность у зарянки вызывает в основном красная грудь сородича, а драка у колюшек возникает как реакция в первую очередь на красное брюхо другого самца. «Усы» самца золотого дятла значат для сородичей больше любого другого его признака и т. д. По-видимому, у таких цветов, форм, звуков, движений только одна функция — вызывать соответствующую реакцию другой особи. Впервые мысль о том, что социальные реакции часто запускаются как будто специально приспособленными для этого стимулами, была четко сформулирована Лоренцем. Такие признаки он назвал релизерами[2]. Лоренц описывает свою концепцию релизеров следующим образом: «Средства, выработавшиеся для посылки ключевых стимулов, могут заключаться в морфологических признаках типа особого цветного узора или специфической структуры или в инстинктивном действии типа принятия особой позы, «танца» и т. п. В большинстве случаев они включают оба компонента, т. е. определенные инстинктивные действия служат для демонстрации узоров или структур, развившихся специально для этих целей. Все эти приспособления, используемые для запуска [82] заданной реакции, я назвал релизерами независимо от того, имеют они зрительную или звуковую природу, являются действием, структурой или цветом».
Данные, собранные на сегодняшний день специалистами, работающими в этой области, по-видимому, в целом подтверждают гипотезу Лоренца. Правда, лишь в нескольких случаях доказана ее полная состоятельность, и впереди еще много неясного, но в общем принцип релизеров представляется весьма полезным для понимания механизмов социального сотрудничества. В приводимом ниже обзоре таких стимулов они будут рассмотрены в зависимости от сенсорной модальности без учета связанной с ними функции.
Обзор релизеров.
Звуки важны для групп с хорошо развитыми органами слуха. Уже говорилось, что самцы многих видов привлекают самок специфическими громкими криками, которые если кажутся человеку приятными, получают название песен. Экспериментальных данных в пользу воздействия песни как релизера мало, и в опытах следовало бы шире использовать множество имеющихся записей птичьих сигналов. Возможно, «блеяние» самца бекаса (рис. 50), треск самца козодоя и барабанная дробь самца дятла выполняют строго определенные функции, но экспериментально это не проверено.
Другая группа животных, в жизни которых важную роль играют крики и «песни»,— лягушки и жабы. Естественно, всем знакомо кваканье самцов травяной лягушки и обыкновенной жабы. В субтропиках и тропиках обитает гораздо больше шумных видов земноводных, причем у некоторых из них настолько мелодичные голоса, что назвать их песней можно с гораздо меньшей натяжкой, чем звуки, издаваемые нашими лягушками. О точной функции этих песен и других криков амфибий известно пока мало. [83]

Рис. 50. «Блеющий» бекас звук создается вибрацией наружных перьев хвоста.
Хотя некоторые экспериментальные данные говорят о том, что стрекотание кобылок и сверчков выполняет в принципе ту же роль, что и птичье пение, значение звуков, издаваемых другими насекомыми, почти совсем не изучено. Ухаживающий самец кузнечика издает серию различных «трелей»; они видоспецифичны и у каждого вида весьма устойчивы. Кроме того, звуковые сигналы свойственны цикадам, муравьям и некоторым другим группам насекомых, но их функции неясны.
Довольно распространены и химические сигналы, действующие на органы обоняния, но и в этом случае их функция понятна лишь в немногих случаях. Уже упоминалось, что самцов бабочек привлекает к самкам запах, а млекопитающие оставляют пахучие метки на границах своей территории. Запах бывает важен и при ухаживании как «соблазняющее» средство, стимулирующее взаимодействие со стороны самки. Такова функция пахучих чешуек самца бабочки сатира. Они собраны узкой полосой на верхней стороне переднего крыла самца. Их щетковидная форма способствует испарению секретов пахучих желез (рис. 51). Кульминация ухаживания самца, стоящего перед самкой,— его «поклон»: расправляя передние крылья, он захватывает между ними антенны самки. В результате терминальные органы обоняния, расположенные на булавовидных расширениях ее антенн, приводятся в соприкосновение с пахучими участками. Самцы с удаленными пахучими чешуйками и с их основаниями, покрытыми шеллаком, добиваются меньшего успеха при ухаживании, чем контрольные самцы с покрытыми шеллаком другими частями крыла.
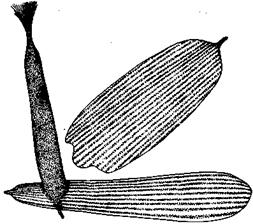
Рис. 51. Две обычные и одна пахучая чешуйки самца сатира Семела.

Рис. 52. Спаривание виноградных улиток. Справа — «любовная стрела». [84]
В социальном сотрудничестве используются и осязательные стимулы. Когда самец колюшки приводит самку к гнезду, она входит в него и готовится метать икру. Однако последнее требует тактильной стимуляции со стороны самца, который «тормошит» партнершу.
Другой пример использования при ухаживании осязательных стимулов — спаривание виноградной улитки (Helix pomatia) (рис. 52). Поскольку эти моллюски — гермафродиты, оба пола ухаживают друг за другом с одинаковой интенсивностью. Церемония состоит из серии поз и движений, завершающейся совокуплением. Шимански сумел спровоцировать полный цикл ухаживания, имитируя стимулы, обычно исходящие от партнера, путем мягких касаний улитки кисточкой. Кульминацией такого «тактильного ухаживания» служит очень мощный стимул: в тело партнера вонзается острое известковое образование — так называемая «любовная стрела», воздействие которой и вызывает собственно совокупление.
Как уже говорилось, угрожающая демонстрация у многих рыб включает в себя тактильные стимулы особого рода, воздействующие на органы боковой линии.
Брачное поведение различных тритонов сопровождается серией сигналов зрительной, осязательной и химической природы. Самец обыкновенного тритона начинает ухаживание, располагаясь перед самкой с поднятым гребнем и повернувшись к ней боком (рис. 53). Затем он совершает внезапный рывок, посылая на партнершу сильный поток воды, который часто отбрасывает ее в сторону. После этого самец располагается головой к самке, направляет хвост вперед параллельно туловищу и, волнообразно его изгибая, гонит на партнершу тихий поток воды, возможно переносящий химический стимулятор (рис. 54). Если та реагирует [85] на это, продвигаясь к самцу, он разворачивается и сначала ползет от нее, спустя некоторое время останавливается, ждет, когда она прикоснется к его хвосту, а после этого откладывает сперматофор, который самка втягивает в свою клоаку. И здесь требуются экспериментальные исследования для проверки очевидного предположения о том, что первое движение тритона — зрительная демонстрация, второе посылает тактильные стимулы, а третье связано с химическими сигналами.
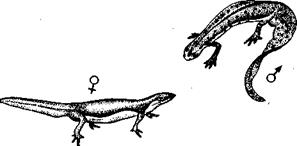
Рис. 53. Фаза зрительной демонстрации у обыкновенного тритона.
Рис. 54. Самец обыкновенного тритона направляет поток воды на самку.
Зрительные релизеры известны относительно хорошо, хотя и в их случаях еще требуется много уточнений. Приведенные выше примеры лишь показывают, что в качестве таких стимулов могут использоваться движение, форма и цвет. У некоторых видов основную роль играет движение, например при различных демонстрациях, связанных с ухаживанием и угрозой у серебристой чайки. В других случаях главное — цвет; достаточно вспомнить о красном брюхе колюшки или красном пятне на нижней челюсти серебристой чайки. Обычно цвет и движение воздействуют вместе, и потом движение всегда удобно для демонстрации особых [86] структур, влияющих на реагента. Приспосабливается ли движение к структуре или структура к движению, или они приспосабливаются друг к другу — проблема эволюционная. Я еще вернусь к ней в гл. 8.
Заключение.
Насколько удалось выяснить к настоящему времени, социальное сотрудничество, по-видимому, зависит в основном от системы релизеров. Тенденция особей подавать такие сигналы — врожденная, тенденция реагировать на них определенными действиями — также. Социальные релизеры, по-видимому, всегда хорошо заметны и относительно просты. Это важно, поскольку из других работ известно, что любые сигналы, «запускающие» врожденные формы поведения, всегда являются простыми «знаковыми стимулами». Следовательно, структуры и элементы поведения, действующие как релизеры, сформировались как особого рода знаковые стимулы. Поскольку одна из функций релизеров — обеспечение репродуктивной изоляции, они видоспецифичны, т. е. отличаются от используемых другими видами. Таким свойством не всегда обладает отдельный релизер, но оно характерно для их последовательности, достигающей высокой специфичности в целом даже при невысокой специфичности каждого ее элемента.
Однако не вся коммуникация основана на релизерах; проблема несколько сложнее. Как уже говорилось, многие социальные животные реагируют на релизеры своего вида, только когда они исходят от определенных особей, известных данному животному «лично». Значит, индивидуальные связи, установившиеся в процессе научения, приводят к тому, что особь реагирует только на сигналы, поступающие от одного или нескольких индивидов; это также реакция на «общевидовые» релизеры, но проявляющаяся лишь после ограничения внимания отдельными сородичами.
Ответ реагента иногда выражается в немедленном и простом движении. Однако часто природа реакции внутренняя. В таком случае сигнал изменяет состояние особи и подготавливает ее к более сложной и разнообразной деятельности.
Таким образом, можно видеть, что функционирование сообщества обеспечивается свойствами его членов. У каждого из них есть тенденция совершать сигнальные движения, запускающие «правильные» реакции у других особей; каждый член сообщества способен специфически воспринимать сигналы своего вида. В этом смысле сообщество определяется индивидами.
Социологи и философы иногда заявляют, что индивиды определяются потребностями сообщества; на первый взгляд это мнение прямо противоположно высказанному выше. Дискуссии по данному вопросу привели к заключению, что в каком-то смысле [87] верны оба положения. Первое правомерно с «физиологической» точки зрения, а второе — с эволюционной. Когда индивиды ведут себя ненормально, сообщество, естественно, страдает, и в этом смысле ясно, что оно определяется составляющими его особями. Однако выживают только группы, образованные нормальными индивидами, поскольку только они способны должным образом размножаться. Значит, результат сотрудничества особей непрерывно проверяется на уровне всей группы, которая, таким образом, самим своим существованием определяет сохранение тех или иных свойств индивидов. Такое рассуждение можно применить и к особи с составляющими ее органами. Особь, естественно, определяется своими органами в том смысле, что неправильное функционирование одного из них угрожает жизни всего организма. Однако результат взаимодействия органов, т.е. сама особь, существует как единое целое, и выживают только экземпляры с правильно функционирующими органами. Таким образом, успех всего организма в конечном итоге определяет свойства его составляющих. [88]
Не нашли, что искали? Воспользуйтесь поиском: