ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Наследование поведения
Согласно гипотезе формативной причинности, наследование поведения зависит от генетической наследственности, а также от морфогенетических полей, которые контролируют развитие нервной системы и всего животного в целом, а также от моторных полей, создаваемых морфическим резонансом от предыдущих подобных животных. В отличие от этого в общепринятой теории считается, что врожденное поведение «программируется» в ДНК.
Экспериментов по наследованию поведения проводилось сравнительно немного, главным образом потому, что их трудно интерпретировать количественно. Тем не менее были сделаны различные попытки такого рода: например, в экспериментах на крысах и мышах поведение «измерялось» скоростью их бега в «колесе», частотой и длительностью их половой активности, интенсивностью дефекации, определяемой как число фекальных шариков, оставляемых на данной площади в единицу времени, способностью к обучению в лабиринте и восприимчивостью к аудиогенным инсультам, вызываемым очень сильными шумами. Наследуемый компонент этих реакций демонстрировался выведением потомства от животных с большим или малым количеством полученных очков: потомство обнаруживало тенденцию к получению очков, близких к тем, которые были у родителей[223]. Проблема с исследованиями такого рода состоит в том, что они дают очень мало сведений о наследовании моделей поведения; более того, результаты трудно интерпретировать, поскольку на них может влиять множество различных факторов. Например, меньшая скорость вращения колеса или пониженная частота спаривания может быть следствием общего снижения тонуса в результате наследуемой метаболической недостаточности.
В некоторых случаях причины генетических изменений поведения исследовались довольно подробно. У маленького круглого червя (нематоды) Caenorhabditis у некоторых мутантов, которые извиваются ненормально, имеются структурные изменения в нервной системе[224]. У дрозофил различные поведенческие мутации, устраняющие нормальный отклик на свет, влияют на фоторецепторы или периферические зрительные нейроны[225]. Известно, что у мышей многие мутации влияют на морфогенез нервной системы, что приводит к дефектам целых участков мозга. У человеческих существ различные врожденные аномалии нервной системы связаны с аномалиями в поведении, например при синдроме Дауна, разновидности монголизма. На поведение могут влиять также наследственные физиологические и биохимические дефекты: например, у человека состояние фенилкетонурии, связанное с умственной неполноценностью, обусловлено недостатком фермента фенилаланингидроксилазы.
Тот факт, что на врожденное поведение влияют генетически обусловленные изменения в структуре и функциях органов чувств, нервной системы и т. д., конечно, не доказывает, что наследование поведения можно объяснить воздействием одних лишь генетических факторов; он показывает только, что для нормального поведения необходимо нормальное тело. Подумайте снова об аналогии с радио: изменения в приемнике влияют на его работу, но это не доказывает, что музыка, которая исходит из громкоговорителя, рождается внутри самого приемника.
В сфере поведения биохимические, физиологические и анатомические изменения могут предотвратить появление зародышевых структур, и, следовательно, целые моторные поля могут быть не в состоянии действовать, или они могут оказывать разные количественные воздействия на движения, контролируемые этими полями. И действительно, исследования в области наследования фиксированных моделей действия показывают, что «нетрудно найти вариации, которые незначительно влияют на выполнение этого действия, но такая модель все же проявляется во вполне узнаваемой форме, если она проявляется вообще»[226].
Наследование моторных полей, возможно, зависит от факторов, которые уже обсуждались в связи с наследованием морфогенетических полей (глава 7). Вообще говоря, у гибридов между двумя породами или видами преобладание моторных полей одного над моторными полями другого, вероятно, зависит от относительной силы морфического резонанса со стороны родительских типов (ср. рис. 19). Если один принадлежит к хорошо установившимся породе или виду, а другой — к относительно новым, с малочисленной предшествующей популяцией, можно ожидать, что моторные поля первого будут доминировать. Но если родительские породы или виды утвердились одинаково хорошо, гибриды будут подпадать под влияние их обоих в одинаковой степени.
И это то, что происходит на самом деле. В некоторых случаях результаты принимают весьма причудливую форму, поскольку модели поведения родительских типов несовместимы друг с другом. Один из примеров дают гибриды, которые получаются при скрещивании двух видов неразлучников (небольших попугаев). Оба родительских вида строят свои гнезда из полосок, которые они вырывают из листьев одинаковым способом; но если один (неразлучник Фишера) несет потом эти полоски к гнезду в клюве, другой (неразлучник персиковоликий) переносит их, засовывая между своими перьями. Гибриды выдергивают полоски из листьев нормально, но затем ведут себя весьма странным образом, иногда засовывая полоски между перьев, иногда перенося их в клюве; но, даже когда они переносят их в клюве, они топорщат перья внизу спины и пытаются спрятать туда свои полоски[227].
11.2. Морфический резонанс и поведение: экспериментальная проверка
В механистической биологии проводится резкое различие между врожденным и обученным поведением: предполагается, что первое «генетически запрограммировано», или «закодировано», в ДНК, тогда как последнее рассматривается как результат физико-химических изменений в нервной системе. Невозможно представить, каким способом эти изменения могут специфически модифицировать ДНК (такое требование выдвигали бы последователи ламаркизма); поэтому считается невозможным, чтобы обученное поведение, приобретенное животным, могло наследоваться его потомством (конечно, исключая «культурное наследование», при котором потомство воспринимает модели поведения от своих родителей).
В противоположность этому, согласно гипотезе формативной причинности, нет качественного различия между врожденным и обученным поведением, поскольку оба зависят от моторных полей, даваемых морфическим резонансом (раздел 10.1). Поэтому эта гипотеза допускает возможную передачу обученного поведения от одного животного к другому и приводит к проверяемым предсказаниям, которые отличаются не только от таковых ортодоксальной теории наследования, но также и от предсказаний ламаркизма.
Рассмотрим следующий эксперимент. Дикие животные помещаются в условия, где они учатся реагировать на данный стимул неким характерным образом. Затем их заставляют повторять эту модель поведения много раз. Согласно гипотезе, новое моторное поле будет усилено морфическим резонансом, который не только приведет к тому, что поведение обученных животных станет все более привычным, но и повлияет так же, хотя и менее специфично, на других животных, подвергаемых тому же стимулу: чем больше будет число животных, научившихся выполнять данное задание в прошлом, тем легче его будет выучить последующим подобным же животным. Поэтому в экспериментах такого типа должна быть возможность наблюдать прогрессивное увеличение скорости обучения не только у животных, происходящих от обученных предков, но также у генетически подобных животных, происходящих от необученных предков. Это предсказание отличается от такового ламаркистской теории, согласно которой только потомки обученных животных должны обучаться быстрее. А по общепринятой теории не должно наблюдаться увеличение скорости обучения потомков как необученных, так и обученных животных.
Сказанное можно суммировать следующим образом: повышенная скорость обучения в последовательных поколениях как тренированных, так и нетренированных линий подтверждала бы гипотезу формативной причинности: увеличение скорости только в тренированных линиях — ламаркизм, а отсутствие такого увеличения в обоих линиях — ортодоксальную теорию.
Эксперименты этого типа фактически уже проводились. Результаты подтверждают гипотезу формативной причинности.
Оригинальный эксперимент был начат в Гарварде в 1920 году У. Мак-Дугаллом, который надеялся провести тщательную проверку возможности наследования приобретенных особенностей поведения, предполагаемых ламаркизмом. Экспериментальными животными были белые крысы уистарской (Wistar) линии, которые -бережно выращивались в лабораторных условиях в течение многих поколений. Их задача состояла в том, чтобы научиться выбираться из специально сконструированного бака с водой, доплывая до одного из двух проходов, из которого можно было выйти наружу. Ложный проход был ярко освещен, тогда как истинный проход не освещался. Если крыса выходила через освещенный проход, она получала удар электрошоком. Два прохода освещались по очереди, один раз — первый, следующий раз — второй. За меру скорости обучения принималось число ошибок, которые допускала крыса, прежде чем она усваивала, что выходить надо через неосвещенный проход:
«Некоторым крысам требовалось целых 330 погружений, причем приблизительно половина из них сопровождалась электрошоком, прежде чем они научались избегать освещенный проход. Во всех случаях процесс обучения внезапно достигал критической точки. В течение длительного времени животное выказывало явное отвращение к освещенному проходу, часто колебалось, прежде чем войти в него, поворачивало назад или отчаянно бросалось внутрь; но, не уловив постоянной связи между ярким светом и шоком, оно продолжало устремляться в этот проход столь же часто, как и в другой. Наконец в обучении наступал момент, когда животные, оказавшись перед освещенным проходом, определенно и решительно поворачивали обратно, искали другой, смутно видимый проход и спокойно вылезали наружу. После достижения этой точки в обучении ни одно животное не делало ошибку, снова выбирая яркий проход, разве что в очень редких случаях» [228].
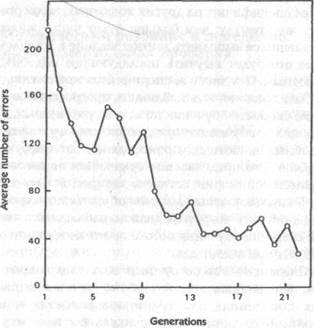
Рис. 28. Среднее число ошибок у последовательных поколений крыс, отобранных в каждом поколении по медленности обучения (данные из McDougall, 1938)
В каждом поколении крысы, от которых должно было быть выведено следующее поколение, отбирались случайным образом до того, как определялась скорость их обучения, тогда как спаривание происходило после их тестирования. Такая процедура была выбрана для того, чтобы избежать любой возможности сознательного или неосознанного отбора в пользу более быстро обучающихся крыс.
Этот эксперимент продолжался на протяжении 32 поколений крыс в течение 15 лет. В согласии с теорией Ламарка в последовательных поколениях крыс наблюдалась заметная тенденция к увеличению скорости обучения. Об этом свидетельствовало среднее число ошибок, которые делали крысы: в первых восьми поколениях оно превышало 56, а во второй, третьей и четвертой группах из восьми поколений — соответственно, 41, 29 и 20[229]. Разница была очевидной не только в количественных данных, но также в фактическом поведении крыс, которые в последующих поколениях становились более осторожными и опытными[230].
Мак-Дугалл предвидел критику того рода, что, несмотря на случайный отбор родителей в каждом поколении, все же мог вкрасться какой-то отбор в пользу более быстро обучающихся крыс. Чтобы проверить такую возможность, он начал новый эксперимент с другой группой крыс, в которой родители действительно отбирались на основе очков, полученных ими в процессе обучения. В одной серии выбирались только быстро, в другой — только медленно обучающиеся крысы. Как и ожидалось, потомство быстро обучающихся крыс обучалось сравнительно быстро, а потомство медленно обучающихся — сравнительно медленно. Однако даже в последней серии обучение у последних поколений заметно улучшалось, несмотря на повторяющийся отбор в пользу медленно обучающихся крыс {рис. 28).
Эти эксперименты проводились весьма тщательно, и критики не могли отвергнуть результаты по причине технических изъянов. Но они привлекли внимание к недостаткам в постановке экспериментов: Мак-Дугаллу не удалось осуществить систематическую проверку изменения скорости обучения крыс, родители которых не проходили обучения.
Один из этих критиков, Ф. А. Е. Крю из Эдинбурга, повторил эксперименты Мак-Дугалла с крысами, происходившими от той же инбридинговой линии[231], используя бак подобной же конструкции. Он включил также параллельную линию «нетренированных» крыс, отдельные из которых тестировались в каждом поколении на скорость обучения, тогда как другие, которые не тестировались, рождали потомство. Через 18 поколений в таком эксперименте Крю не обнаружил систематического изменения в скорости обучения ни в тренированной, ни в нетренированной линиях[232]. Вначале казалось, что это порождает серьезные сомнения в результатах Мак-Дугалла. Однако эти две группы результатов нельзя было непосредственно сравнивать в силу трех обстоятельств. Во-первых, по какой-то причине крысы гораздо легче обучались в опытах Крю, чем в более ранних поколениях Мак-Дугалла. Этот эффект был столь сильно выражен, что значительное число крыс как в тренированных, так и в нетренированных линиях «решали» задачу немедленно, не получая ни единого шока! Средние значения очков у крыс в опытах Крю с самого начала были примерно такими же, как у крыс Мак-Дугалла через более чем 30 поколений тренировки. Ни Крю, ни Мак-Дугалл не могли дать удовлетворительного объяснению этому несоответствию. Но, как указывал Мак-Дугалл, поскольку цель исследования состояла в том, чтобы выявить любые воздействия тренировки на последующие поколения, эксперимент, в котором некоторые крысы не тренировались вообще, а многие другие — очень мало, нельзя было рассматривать как способный продемонстрировать такое воздействие[233]. Во-вторых, в результатах Крю обнаруживались большие и кажущиеся случайными флуктуации от поколения к поколению, гораздо большие, чем флуктуации в результатах Мак-Дугалла, и эти большие флуктуации вполне могли скрыть любую тенденцию к улучшению очков в более поздних поколениях. В-третьих, Крю выбрал путь очень жесткого инбридинга (скрещивания очень близких родственников), скрещивая в каждом поколении братьев с сестрами. Он не ожидал, что это даст неблагоприятный эффект, поскольку крысы происходили от одной главней инбридинговой семьи, с которой все начиналось:
«Даже история моей главной семьи выглядит как эксперимент по инбридингу. Имеются широкая основа семейных линий и узкий кончик двух оставшихся линий. Скорость воспроизведения падает, и угасает одна линия за другой» [234].
Даже в выживающих линиях многие животные рождались со столь серьезными аномалиями, что их приходилось отбрасывать. Вредные последствия такого жесткого инбридинга вполне могли маскировать любую тенденцию к увеличению скорости обучения. В целом эти дефекты эксперимента Крю означают, что его результаты можно рассматривать только как предварительные, и сам он фактически признавал, что вопрос остается открытым[235].
К счастью, эта история здесь не кончается. Эксперимент был снова проведен У. Е. Эгером и его коллегами в Мельбурне с использованием методов без тех недостатков, которые были у Крю. На протяжении 20 лет они измеряли скорости обучения тренированных и нетренированных линий в 50 последовательных поколениях. В согласии с данными Мак-Дугалла они обнаружили, что у крыс тренированной линии существует явная тенденция к более быстрому обучению в последующих поколениях. Но точно такая же тенденция была обнаружена и в нетренированной линии [236].
Можно удивляться, почему же Мак-Дугалл не наблюдал подобный эффект на своих нетренированных линиях? Ответ состоит в том, что он его наблюдал.
Несмотря на то что он тестировал контрольных крыс, происходивших от исходной главной семьи лишь изредка, он заметил «неприятный факт, что группы контрольных крыс, произведенных от этой семьи в 1926, 1927, 1930 и 1932 годах, демонстрировали уменьшение среднего числа ошибок с 1927-го по 1932 год». Он думал, что этот результат мог быть случайным, но добавил следующее:
«Возможно, что выпадение по среднему числу ошибок периода с 1927-го по 1932 год выражает реальное изменение конституции всей семьи, ее улучшение (относительно этой определенной способности), природу которого я не могу предположить» [237].
С публикацией окончательного отчета группы Эгера в 1954 году закончилась затянувшаяся полемика по поводу «ламаркистского эксперимента Мак-Дугалла». Одинаковое улучшение результатов как в тренированной, так и в нетренированной линиях исключало интерпретацию в духе ламаркизма. Вывод Мак-Дугалла был отвергнут, и это, казалось, исчерпывало вопрос. С другой стороны, его результаты получили подтверждение.
Эти результаты казались совершенно необъяснимыми; они были бессмысленными с точки зрения какой-либо из действующих концепций и никогда не были доведены до конца. Но они вполне осмысленны в свете гипотезы формативной причинности. Конечно, сами по себе они не могут доказать справедливость этой гипотезы; всегда можно предложить другие объяснения, например то, что последовательные поколения крыс становились все более разумными по неизвестной причине, не связанной с их тренировкой[238].
В будущих экспериментах наиболее прямым способом проверки действия морфического резонанса, вероятно, будет такой, в котором большое число крыс (или любых других животных) обучается новой задаче в одном месте, а затем такие же крысы учатся выполнять ту же задачу в другом месте, за сотни миль от первого, и будет выясняться, есть ли увеличение в скорости обучения у этих последних. Начальная скорость обучения в обеих группах должна быть более или менее одинакова. Тогда, согласно гипотезе формативной причинности, скорость обучения должна прогрессивно возрастать в том месте, где тренируется большое число животных; и такое же увеличение скорости должно обнаруживаться также в другом месте, даже несмотря на то что здесь число испытуемых крыс очень мало. Очевидно, следует принять меры предосторожности, чтобы избежать любых возможных сознательных или неосознанных предубеждений со стороны экспериментаторов. Один из способов может состоять в том, чтобы во втором месте тестировать скорости обучения крыс, давая им несколько разных задач с правильным интервалом, например раз в месяц. Затем в первом месте выбирается наугад из этого набора определенная задача, которой обучаются тысячи крыс. Более того, время начала обучения также будет выбрано наугад, например четыре месяца спустя после начала выполнения набора задач во втором месте. Экспериментаторы во втором месте не будут знать ни того, какая задача была выбрана, ни того, когда началось обучение ей в первом месте. Если при этих условиях во втором месте будет обнаружено заметное увеличение в скорости обучения данной задаче после того, как обучение началось в первом месте, этот результат будет весомым подтверждением справедливости гипотезы формативной причинности.
Эффект такого типа вполне мог иметь место, когда группы Крю и Эгера повторяли эксперимент Мак-Дугалла. В обоих случаях их крысы с самого начала обучались своей задаче значительно быстрее, чем это делали крысы Мак-Дугалла, когда он впервые начал свой эксперимент[239].
Если предложенный выше эксперимент действительно был бы осуществлен и дал бы положительные результаты, он не мог бы быть полностью воспроизводим по самой своей природе, ибо при попытках его повторить на крыс должен будет действовать морфический резонанс от крыс в первоначальном эксперименте. Чтобы продемонстрировать тот же эффект снова и снова, было бы необходимо изменить либо задачу, либо вид, тестируемый в каждом эксперименте.
Эволюция поведения
Хотя палеонтология дает прямые сведения о структуре животных, живших в прошлом, она практически ничего не сообщает об их поведении. Вследствие этого большинство представлений об эволюции поведения может основываться не на свидетельствах, пришедших из прошлого, но лишь на сравнениях между видами, существующими в настоящем. Так, например, теории эволюции общественного поведения пчел могут строиться путем сравнения ныне существующих общественных видов с обособленными и колониальными видами, предположительно более примитивными. Но сколь бы разумными ни казались эти теории, они всегда будут не более чем спекулятивными[240]. Более того, теории эволюции поведения зависят от предположений относительно способа наследования поведения, поскольку на сегодняшний день об этом фактически известно очень немного.
Механистическая, или неодарвинистская, теория предполагает, что врожденное поведение «программируется», или «кодируется», в ДНК и что новые типы поведения обусловлены случайными мутациями. Затем естественный отбор благоприятствует полезным мутантам — так развиваются инстинкты. Предполагается, что случайные мутации дают животным также способности к определенным видам обучения. Тогда животные, у которых выживаемость и воспроизводство себе подобных выигрывают от этих способностей, переживают в результате естественного отбора. Таким образом развиваются способности к обучению. Даже то, что обученное поведение имеет тенденцию становиться врожденным, можно приписать случайным мутациям, основываясь на гипотетическом эффекте Болдуина: животные могут реагировать на новые ситуации, обучаясь соответствующим способам поведения; случайные мутации, которые вызвали к жизни эти типы поведения без необходимости обучения, будут закрепляться естественным отбором; таким образом, поведение, которое вначале было приобретенным, может стать врожденным не из-за наследования приобретенных признаков, а потому что случайно произошли соответствующие мутации.
Кажется, практически не существует предела тому, что может быть объяснено с помощью благоприятных случайных мутаций, которые изменяют «генетическое программирование» поведения. Затем эти неодарвинистские теории могут быть разработаны в математической форме посредством вычислений, основанных на формулах теоретической популяционной генетики[241]. Но поскольку эти спекуляции невозможно проверить, они не имеют независимой научной ценности; они лишь развивают механистические допущения, из которых проистекли.
Гипотеза формативной причинности приводит к совершенно отличным интерпретациям эволюции поведения. Можно ожидать, что естественный отбор приводит к изменениям «генных пулов» популяций до той степени, в какой генетические изменения влияют на поведение. Но сами специфические модели поведения зависят от наследования моторных полей путем морфического резонанса. Чем больше повторяется данная модель поведения, тем сильнее становится этот резонанс. Таким образом, повторение инстинктивного поведения будет все более и более закреплять инстинкты. С другой стороны, если тип поведения варьируется от индивидуума к индивидууму, морфический резонанс не даст четко оформленных хреод; следовательно, поведение будет менее стереотипным. И чем больше разнообразия в поведении, тем больше будет диапазон вариаций в будущих поколениях. Этот тип эволюции в направлении, допускающем появление разума, до некоторой степени осуществился у птиц, еще более у млекопитающих и более всего у человека.
В некоторых случаях полуобученное поведение должно было развиться из основы, в которой оно было полностью инстинктивным. Одним из путей такой эволюции могла быть гибридизация пород с различными хреодами, вызывающая появление составных (композитных) моторных полей с большим диапазоном индивидуальных вариаций.
В других случаях полуинстинктивное поведение могло эволюционировать из поведения, которое изначально было обученным, в результате частых повторений. Рассмотрим, например, поведение различных пород собак. Овчарки в течение многих поколений обучались и отбирались по способности загонять овец, охотничьи собаки — по способности находить и подавать дичь, пойнтеры — делать стойку, гончие на лис — преследовать лис и так далее. Собаки часто проявляют врожденное стремление к поведению, характерному для их породы, даже раньше, чем их начинают обучать[242]. Возможно, такие тенденции не столь сильны, чтобы их можно было назвать инстинктами, но они достаточно сильны, чтобы показать, что между инстинктом и наследственной предрасположенностью к обучению определенным типам поведения существует различие лишь в степени. Конечно, породы собак развивались в условиях, скорее, искусственного, чем естественного отбора, но кажется вероятным, что в обоих случаях действуют одинаковые принципы.
Хотя относительно легко представить, как некоторые типы инстинктивного поведения могли развиться путем повторения обученного поведения в одном поколении за другим, это не может легко объяснить эволюцию всех типов инстинкта, особенно у животных с очень ограниченной способностью к обучению. Быть может, некоторые новые инстинкты появились из новых изменений и сочетаний инстинктов, существовавших ранее; одним из возможных путей здесь могла быть гибридизация пород или видов с различными моделями поведения. Другой путь появления новых комбинаций — включение в старые модели «сдвиговых видов деятельности» (displacement activities), то есть кажущихся неадекватными действий животных, «раздираемых» противоречащими друг другу инстинктами. Таким путем вполне могли возникнуть некоторые виды ритуалов ухаживания[243]. Можно представить также, что мутации или необычные условия среды могут сделать животное способным «настраиваться» на моторные хреоды других видов (ср. раздел 8.6).
Но в дополнение к рекомбинации существующих хреод должен быть также способ, посредством которого совершенно новые моторные поля возникают у животных, поведение которых почти полностью инстинктивно. Новые модели поведения могут появляться только в том случае, если привычное повторение наследственного поведения было заблокировано в результате либо изменения окружения, либо мутации, изменившей нормальную физиологию или морфогенез животного. В большинстве таких случаев животное будет действовать некоординированно и неэффективно, но иногда при этом может возникать и новое моторное поле. И когда бы новое поле ни появилось впервые, должен произойти «скачок», который не может быть полностью объяснен действием предшествовавших энергетических или формативных причин (разделы 5.1, 8.7).
Если модель поведения, обусловленная новым моторным полем, нарушает способность животных к выживанию и воспроизведению, она не станет повторяться очень часто, ибо животные, которые повторяют такое поведение, будут устранены (элиминированы) действием естественного отбора. Но если эта модель поведения помогает животным выживать и воспроизводиться, она будет повторяться часто и поэтому будет все более усиливаться морфическим резонансом. Таким образом, моторному полю будет благоприятствовать естественный отбор.
Поведение человека
Высшие животные часто ведут себя более гибко, чем низшие животные. Однако эта гибкость ограничена ранними стадиями поведенческого ряда, и особенно начальной фазой, обусловленной потребностью в пище; более поздние стадии, и в частности акт осуществления, протекают стереотипным образом как фиксированные модели поведения (раздел 10.1).
В «ландшафтной» модели главное моторное поле может быть представлено широкой долиной, которая затем сужается, края ее становятся все более крутыми и, наконец, она заканчивается глубоким каньоном {рис. 27 В). Широкая долина соответствует фазе питания, на которой можно использовать множество альтернативных путей; затем эти пути сливаются, так как они «втягиваются в воронку» по направлению к сильно канализованной хреоде конечного акта.
В человеческом поведении круг возможностей, используемых для достижения поведенческих целей, гораздо шире, чем у любого другого вида, но и здесь, по-видимому, применимы те же принципы: под воздействием моторных полей высших уровней модели поведения «втягиваются в воронку», ведущую к стереотипным актам осуществления, которые обычно являются врожденными. Например, люди получают пищу всевозможными способами: либо непосредственно — охотясь, ловя рыбу, пася скот или возделывая поле, либо косвенно — выполняя различные работы. Затем еду готовят множеством различных способов и переносят в рот также по-разному, например рукой, палочками для еды или ложкой. Но способы жевания еды различаются весьма незначительно, а завершающий акт всего моторного поля питания, глотание, одинаков у всех людей. Подобно этому, в поведении, управляемым моторным полем репродукции, методы ухаживания и системы вступления в брак широко варьируются, но завершающий акт совокупления, к которому они ведут, более или менее стереотипен. У мужчин конечная фиксированная модель поведения, извержение семени (эякуляция), протекает автоматически и фактически является врожденной.
Таким образом, большое разнообразие моделей поведения человека обычно направлено к ограниченному числу целей, задаваемых моторными полями, унаследованными от предшествовавших членов вида путем морфического резонанса; в общем случае эти цели связаны с развитием, сохранением или воспроизведением индивидуума или социальной группы. Даже игра и исследовательская деятельность, не направленные непосредственно к этим целям, часто помогают достичь их позднее, как они делают это у других видов. Поскольку ни игра, ни «обобщенное исследовательское питательное поведение» в отсутствие немедленной награды не ограничены только человеком: например, крысы исследуют свое окружение и объекты в нем, даже когда они сыты[244].
Однако не вся человеческая деятельность подчинена моторным полям, которые канализируют ее в направлении биологических или социальных целей; некоторые виды деятельности явно направлены к трансцендентным целям. Этот тип поведения в наиболее чистом виде представлен жизнью святых. Но ясно, что преобладающая часть поведения большинства человеческих существ не имеет такой трансцендентной направленности.
Несмотря на то что диапазон различий человеческого поведения очень широк, когда рассматривается вид в целом, в любом данном обществе деятельность индивидуумов лежит в пределах ограниченного числа стандартных моделей. Люди обычно повторяют характерным образом структурированные виды деятельности, которые уже выполнялись снова и снова многими поколениями их предшественников. Эти виды включают: знание определенного языка; двигательные навыки, связанные с охотой, сельским хозяйством, ткачеством, изготовлением инструментов, приготовлением пищи и так далее; пение и танцы; а также типы поведения, специфичные для определенных социальных ролей.
Все модели деятельности, характерные для данной культуры, можно рассматривать как хреоды[245]. Чем чаще они повторяются, тем более они будут стабилизированы. Но из-за приводящего в замешательство изобилия таких специфичных для культуры наборов хреод, каждая из которых потенциально способна канализировать движения любого человеческого существа, сам по себе морфический резонанс не может ввести индивидуума скорее в один набор хреод, чем в другой. Так что ни одна из этих моделей поведения не проявляется спонтанно: все их нужно узнать. Индивидуум побуждается (инициируется) другими членами общества к принятию определенных моделей поведения. Затем, когда процесс обучения начинается, обычно путем подражания осуществление определенной модели движения приводит человека в состояние морфического резонанса со всеми, кто следовал этой модели в прошлом. Следовательно, обучение облегчается, когда индивидуум «настраивается» на специфические хреоды.
Процессы инициации и в самом деле традиционно понимаются примерно в таких же терминах, какие упомянуты выше. Предполагается, что человек входит в состояния или способы существования, которые предшествуют ему и представляют своего рода трансперсональную реальность.
Облегчение обучения посредством морфического резонанса было бы трудно продемонстрировать в случае давно установившихся моделей поведения, но изменение в скорости обучения должно обнаруживаться легче при двигательных моделях недавнего происхождения. Так, например, в настоящем столетии должно было становиться все легче и легче обучаться езде на велосипеде, вождению автомобиля, игре на фортепиано или работе на пишущей машинке, благодаря кумулятивному морфическому резонансу от большого числа людей, которые уже приобрели эти навыки. Однако, даже если бы надежные количественные данные показали, что скорости обучения действительно возросли, интерпретировать эти данные было бы сложно вследствие возможного влияния других факторов, наподобие усовершенствования конструкции машины, улучшения методов обучения и более высоких побуждений к обучению. Но при специально поставленных экспериментах, в которых приняты меры к тому, чтобы эти факторы сохранялись постоянными, было бы вполне возможно получить убедительные свидетельства существования предсказываемого эффекта.
Гипотеза формативной причинности применима ко всем аспектам поведения человека, в которых повторяются определенные модели движения. Но она не может объяснить происхождение этих моделей. Здесь, как и в других случаях, проблема творческой деятельности (creativity) лежит вне пределов естественной науки, и ответ может быть дан лишь с позиций метафизики (ср. разделы 5.1, 8.7 и 11.3).
Глава 12
Четыре возможных вывода
Не нашли, что искали? Воспользуйтесь поиском: