ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Функционирование ниши стволовых клеток АМП.
Основным вопросом функционирования меристем является поддержание баланса между делением клеток и их дифференцировкой в функционировании стволовых клеток ЦЗ. В случае деления стволовых клеток можно наблюдать функциональную асимметрию - одна дочерняя клетка остается стволовой, а другая детерминирована на выполнение определенной функции. Регуляция этого процесса может иметь внутреннюю причину, присущую стволовым клеткам, либо зависеть от неоднородного окружения стволовой клетки. В многоклеточном организме, более правдоподобным кажется механизм, в котором значение имеет именно локализация, нежели автономная работа индивидуальной клетки. Например, в АМП стволовые клетки объединены в группу (ЦЗ), функционирующую, как единое целое, в ответ на сигналы окружающей среды и самого организма, к тому же, деления стволовой клетки не всегда асимметричны - часто стволовая клетка образует две стволовые или две дифференцированные дочерние. Структуры, аналогичные меристемам растений у различных организмов носят название «ниш стволовых клеток» []. Под нишей подразумевается подмножество клеток ткани и внеклеточного вещества, которые могут неограниченно долго «предоставлять убежище» одной или более стволовым клеткам, контролируя процессы их самовоспроизведения и образования дочерних клеток. При этом, отличительной чертой стволовой клетки будет являться именно контакт с клетками ниши, который осуществляется физически- через непосредственный контакт с клетками ниши, или дистантно, через действие морфогенов, распространяющихся от клеток ниши.
Функциональная асимметрия делений стволовых клеток животных обеспечивается непосредственным контактом клеток со специфическим микроокружением (например, с базальной мембраной). В меристемах растений стволовые клетки непосредственно контактируют с внешней средой и не имеют специфического окружения, которое определяло бы их стволовость. Тем ни менее, в генетике растений давно описан механизм в котором размеры ЦЗ регулируются белковым продуктом, экспрессирующемся в нижележащем домене, ОЦ (Sharma V.K. et al., 2003). Этот механизм является в некоторой степени доказательством организации меристем по типу ниши стволовых клеток, в котором регуляция делений стволовых клеток происходит извне. Ключевым геном регуляторного контура, лежащего в основе этого механизма, является ген CLV3, в норме транскрибирующийся только в двух поверхностных слоях ЦЗ и в незначительном числе клеток, относящихся одновременно к ЦЗ и поверхностному слою корпуса. Белок CLV3 выходит во внеклеточный апопласт и мигрирует в пределах АМП. В корпусе экспрессируется рецептор CLV1, который, при наличии сигнала из ЦЗ в виде лиганда CLV3, формирует активный комплекс CLV1/CLV2/KAPP/Rop. (Schoof H. et al., 2000) (Brand et al., 2000) (Stone et al., 1998). Результатом передачи сигнала является подавление этим белковым комплексом экспрессии WUS в тех клетках, куда достигает сигнал CLV3 (негативная связь). В остальных клетках корпуса образования CLV1/CLV2/KAPP/Rop комплекса и, соответственно, подавления транскрипции WUS не происходит. Эти клетки и образуют ОЦ, принадлежность клеток к которому определяется целиком и полностью наличием в них экспрессирующегося гена WUS (описанный механизм не объясняет позиционирования ОЦ в ограниченном количестве клеток под ЦЗ) (Schoof H. et al., 2000) (Sharma et al., 2003). WUS, в свою очередь, увеличивает экспрессию CLV3 в вышележащей ЦЗ (положительная регуляторная связь). При уменьшении количества WUS-экспрессирующих клеток в ОЦ, активация CLV3 происходит в меньшей степени и ЦЗ, экспрессирующая CLV3, уменьшается, что приводит к увеличению числа WUS-экспрессирующих клеток и росту размеров ОЦ. И, наоборот, при увеличении размеров ЦЗ возрастает количество CLV3 в межклеточном пространстве, что влияет на снижение количества клеток в ОЦ в результате подавления экспрессии WUS, меньшая WUS-экспрессирующая зона приводит к уменьшению размеров ЦЗ. Таким образом, имеет место пространственно разобщенный регуляторный контур (Sharma et al., 2003). Такое динамическое равновесие нарушается при мутациях генов WUS и CLV3. При мутации clv3 в АМП увеличивается как ОЦ, так и размеры ЦЗ (Brand et al., 2000). В wus мутантах АМП практически не имеет ни ОЦ, ни ЦЗ (Long et al., 1998). Механизм этого регуляторного контура не до конца понятен, очевидно, существует большое количество дополнительных факторов и регуляторных связей, обеспечивающих его работу. Так, есть данные о миРНК регуляции описанного механизма. Так, при увеличении экспрессии miR166g, домены экспрессии генов WUS и CLV3 в АМП распространяются латерально, а экспрессия WUS увеличена в 12 раз (Williams et al., 2005).
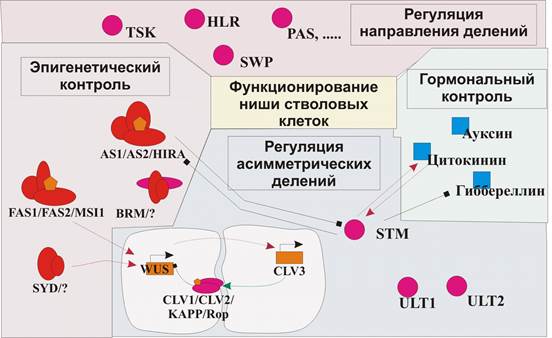
Рис. 2. Регуляторные механизмы, вовлеченные в функционирование ниши стволовых клеток в центральной зоне АМП.
Интересно, что по аналогии с ОЦ в АМП, в корневой меристеме за поддержание пула стволовых клеток отвечает так называемый, покоящийся центр. Покоящийся центр, как и ОЦ, характеризуется экспрессией гена WUS, которая, однако, ниже чем экспрессия WUS в АМП (Hamada S. et al., 2000). При повышении уровня экспрессии WUS в корне, корневая меристема формирует побег (Gallois JL et al., 2004), (Kamiya N. et al., 2003). С другой стороны, в мутантах TPL гена наблюдали обратный процесс: формирование корня из АМП (Long et al., 2002).
Для функционирования ниши стволовых клеток особое значение имеет структура ткани. Нарушения порядка и направления делений клеток в ней приводит и к нарушениям постоянства размеров пула стволовых клеток. Описано большое количество мутаций, в которых побочным эффектом дезорганизации структуры меристемы является увеличение пула стволовых клеток, как правило, за счет нарушения базовых процессов (деградации, формирование веретена деления и прочих). Например, мутация гена HLR, кодирующего субъединицу 26S протеосомы, вызывает дезорганизованность обеих меристем и, как результат, их пониженную активность. В hlr мутантах нарушена экспрессия основных генов, необходимых для функционирования как корневой меристемы так и АМП, в том числе WUS, это говорит о принципиальном значении программного протеолиза для функционирования меристем (Ueda et al., 2004). Сходные аномалии наблюдаются при мутации гена TSK, которая так же приводит к дезорганизации строения обеих меристем. В АМП при этом образуется несколько ОЦ, а в покоящемся центре корня перестает экспрессироваться ключевой ген SCR (Suzuki et al., 2004). Нарушение эпигенетического контроля развития также вызывает целый комплекс аномалий, среди которых дезорганизованная и увеличенная меристема. FAS1, FAS2 и AtMSI1, кодирующие субъединицы хроматин-связывающего комплекса, а также ген SYD необходимы для нормального функционирования АМП. Мутанты этих генов имеют дезорганизованную, увеличенную в размерах АМП (Kaya et al., 2001) (Kwon et al., 2005). Показано, что в мутантах нарушена пространственная экспрессия WUS и CLV3. Мутации по другим факторам ремоделлинга хроматина- AtBRM, AtCAP1 и AtCAP2 также вызывают нарушение стабильности стволового запаса АМП и ее дезорганизованность (Farrona et al., 2004) (Siddiqui et al., 2003).
Помимо CLV-WUS регуляторного контура, изучен и другой, независимый механизм поддержания размеров ЦЗ. Он осуществляется через гомеобоксный ген STM, необходимый как для образования АМП в эмбриогенезе, так и дальнейшего ее развития. В зависимости от локализации мутации в белке STM, растения или не формируют АМП вовсе (stm-5), или имеют АМП меньших размеров (stm-2), или формируют ее уже после прорастания (stm-1, stm-2) (Barton at al., 1993) (Long et al., 1998). В мутантах stm-5 на месте клеток ЦЗ находятся частично дифференцированные клетки, экспрессирующие белки AS1 и AS2. В норме STM подавляет экспрессию AS1 и AS2 и через эту регуляторную связь, опосредует свое влияние на дифференцировку клеток в АМП (Gallois et al., 2002) (Byrne et al., 2000) (Byrne et al., 2002). Показано влияние STM на клеточный цикл через гормон цитокинин. Цитокинины характеризуются способностью вызывать деление клеток и индуцировать дифференцировку побегов. Мутации, нарушающие биогенез цитокинина или передачу цитокининового сигнала, характеризуются ранним прорастанием зародыша и разрастанием АМП, которая формирует большое количество листообразных структур (Hamant et al., 2002) (Rupp et al., 1999). Цитокинин активирует экспрессию STM, KNAT1 и KNAT2 генов. В свою очередь, активация STM повышает уровень мРНК генов, участвующих в биосинтезе цитокинина и передаче цитокининового сигнала, в частности, ARR5. За счет этого уровень цитокинина значительно повышается (Yanai O. et al., 2005) (Jasinski S. et al., 2005). Еще одним механизмом действия STM в АМП является репрессия этим белком ферментов биосинтеза гибереллина, а именно GA5. Показано, что усиленное действие гибереллина, в spy мутантах, губительна для развития АМП: в таких растениях рост растения заканчивается преждевременно, еще до формирования листьев (Hay et al., 2002). Таким образом, предполагается, что STM обеспечивает нормальное функционирование АМП за счет поддержания определенного уровня активности гормонов: одновременной активации цитокининового сигнала и подавления биосинтеза гибереллинов (Jasinski S. et al., 2005).
Недавно описан еще один независимый механизм поддержания баланса стволовых клеток ортологичными генами ULT1 и ULT2. В мутантах ult1 размеры АМП увеличены на стадии цветения, мутация ult2 не имеет какого либо фенотипического проявления в развитии АМП, в то время как двойные мутанты ult1 ult2 приводят к терминации развития АМП вскоре после прорастания. Мутация ULT1 гена частично восстанавливает нормальное функционирование АМП в wus и stm мутантах на вегетативной стадии (Carles et al., 2004).
Не нашли, что искали? Воспользуйтесь поиском: