ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Часть III ЭВОЛЮЦИЯ ПСИХИКИ. 5 страница
В этом примере обращает на себя внимание строгая фиксированность последовательности сигнальных поз и телодвижений, что и характерно для ритуализованного поведения. Как мы видим, при ритуализации двигательные акты всегда носят условный характер: выполняемые животным движения несут лишь сигнальную, но не непосредственно «механическую» функцию. Именно на этой основе зародился язык поз, телодвижений и звуков у животных во всем своем удивительном многообразии. При этом в условности сигналов находит непосредственное воплощение психический компонент общения.
Наиболее сложные системы общения присущи, конечно, насекомым, живущим большими семьями. Это относится опять-таки в первую очередь к медоносной пчеле и муравьям. Дифференцированные и многообразные взаимоотношения между членами пчелиной семьи, опирающиеся на сложные формы коммуникации, являются уникальными в животном мире. По поводу этого сообщества высказывались самые различные мнения начиная от антропоморфичного толкования ее как копии человеческого общества или государства (сохранилось неудачное, вводящее в заблуждение название «общественные насекомые») до представления о некоей сумме независимо действующих «рефлекторных автоматов». Вместе с тем оказалось (исследования К.П.Трубецкой), что пчелиная семья представляет собой объединение особей, поведение которых носит самостоятельный, но не независимый характер. Самостоятельность поведения отдельных пчел проявляется прежде всего в резко выраженных индивидуальных различиях: предпочтение отдельных видов ульевых работ, различия в сроках и характере перехода от ульевой деятельности к летной и др. Большую сложность в количественном и качественном аспектах достигают взаимоотношения между обитателями улья, в результате чего функциональная настроенность одних пчел может оказать влияние на других. Рабочие особи с их повышенным уровнем нервной деятельности часто сообщают возбуждение другим, менее активным рабочим пчелам.
Территориальное поведение.
Уже у кольчатых червей и низших моллюсков наблюдаются первые проявления территориального поведения, что выражается в приуроченности жизнедеятельности к постоянно занимаемому участку, на котором располагается и убежище («дом») животного. Правда, выполнение жизненных функций (или хотя бы части из них) на определенном участке составляет лишь часть территориального поведения, равно как и наличие «дома» само по себе еще не является критерием последнего. При полноценном территориальном поведении, встречающемся лишь на уровне перцептивной психики, животное-хозяин своим видом, голосом или запахом дает знать сородичам о своем присутствии на занятом участке. Важную роль играет при этом «маркировка местности», т.е. нанесение пахучих или иных меток на разные объекты, особенно на периферии участка.
Такими метками могут быть испражнения или выделения кожных желез, а также оптические знаки — содранная со стволов деревьев кора, вытоптанная трава и т.п.
Весьма существенным признаком территориального поведения является активная защита участка хозяином. Тинберген придает этому критерию наряду с привязанностью к местности особое значение. Враждебное отношение («агрессивность») проявляется хозяином по отношению к любому представителю того же вида, особенно того же пола.
Часто такое поведение приурочено лишь к периоду размножения. Так, например, древесный таракан (Cryptocereus punctulatus), устраивающий гнездовую камеру в гнилой древесине, весьма интенсивно (и обычно успешно) защищает место, где находится эта камера, от вторжения других самцов. В случае поражения гнездовую камеру, где находится и самка, занимает победитель.
В высокоразвитых формах территориальное поведение, как показал немецкий этолог А.Хеймер, встречается у стрекоз. Это интересно уже потому, что стрекозы относятся к самым древним насекомым и, появившись в палеозое, ожили до наших дней, не претерпев за эти 50 миллионов лет существенных морфологических изменений. Вместе с тем, как показывает Хеймер, архаичность строения сочетается у них с высокоразвитым поведением, причем не только в сфере территориальности. Этот, казалось бы, парадоксальный факт сохранения древнейших признаков строения при наличии весьма прогрессивных форм поведения вновь подтверждает общее правило несоответствия морфологических и поведенческих признаков, если иметь в виду морфофункциональные отношения на уровне целого организма.
Что же касается территориального поведения стрекоз (речь идет конкретно о равнокрылых стрекозах-красотках Zygoptera), то половозрелые самцы отыскивают места, пригодные для постоянного пребывания, которые и становятся их индивидуальными участками. Эти участки маркируются оптически, а именно путем ежедневных многократных облетов, и энергично обороняются от самцов-сородичей. Внутри индивидуального участка имеются основные и дополнительные места отдыха, а также зона для откладки яиц, границы которой маркируются упомянутым образом. В эту зону самец приводит самку посредством совместно выполняемого с ней «танца» над поверхностью воды. Кроме того, самец «ухаживает» за самкой, выполняя на своей территории особый ритуализо-ванный брачный полет, а во время откладки яиц он охраняет и сторожит самку или соответственно несколько самок, ибо один самец нередко спаривается подряд с несколькими самками. Охрана проявляется в том, что самец отгоняет других мужских особей, пытающихся спариваться с «его» самками, что мешает откладыванию яиц, сторожевая же функция состоит в том, что при попытке самок прервать это занятие самец заставляет их вернуться и возобновить его. Добавим еще, что территориальная борьба самцов выполняется в высокоритуализованных формах (подлинная борьба отсутствует!). В целом все поведение самцов (как и самок) отличается большой пластичностью.
Таким образом, у данных видов стрекоз существует вполне развитое территориальное поведение: занимаются индивидуальные участки, в которых выделяются функциональные зоны (отдыха, размножения), на этих участках выполняются все жизненные функции (за исключением сна: вечером самцы собираются в особых местах ночевок), хозяин дает знать о своем присутствии, маркирует и активно защищает участок.
Интересные особенности территориального поведения муравьев были выявлены советским энтомологом А.А.Захаровым. Оказалось, что у муравьев существуют два основных типа использования кормовых участков: совместное использование угодий несколькими семьями и использование кормового участка населением лишь одного гнезда. При этом выявилась прямая связь между плотностью муравьев на участке и «агрессивностью»: у видов с низкой плотностью участки не охраняются (кроме пригнездовой зоны), при высокой же плотности на кормовых участках появляются охраняемые территории, а между ними «нейтральные зоны» (рис. 40). Как показал другой советский исследователь, Г.М.Длусский, муравьи того же вида, принадлежащие к другим семьям, равно как и представители других видов, в пределы этих территорий не допускаются.

|
Рис. 40. Территориальное поведение муравьев (Acantholepis melanogaster). Территории трех семей с нейтральными зонами между ними. Точки — гнезда; крестики — колонии тлей, посещаемые муравьями; замкнутые кривые — кустарник; пунктирные линии — основные направления поиска и транспортировки пищи (по Захарову)
Наибольшую сложность территориальное поведение достигло у рыжих лесных муравьев, у которых наблюдается и наибольшая плотность особей на кормовых участках. Охраняемые территории этих муравьев достигают значительно большей величины, чем у других видов. Центральными элементами такой территории являются муравьиные тропы (постоянные кормовые дороги), длина которых нередко превышает 100 метров, а протяженность всей сети троп — 1 километр. Каждой дорогой пользуется лишь определенная группа муравьев («колонна»), занимающая в муравейнике определенный сектор., примыкающий к «своей» дороге. Территория всей семьи подразделяется дорогами на отдельные части, составляющие территории отдельных колонн. Между ними имеются такие же нейтральные пространства, как и между территориями семей, хотя и не столь выраженные, как в последнем случае. Границы территории маркированы и препятствуют «рассеиванию» муравьев. Муравьи двигаются по территории не хаотично, а в соответствии со структурой территории: в центральной и средней ее частях - по кратчайшему пути от дорог или муравейника, на периферии же — параллельно границам территории. Таким образом, траектория движения каждого муравья детерминируется местом нахождения его на территории (исследования И.В.Стебаева). Наибольшую охотничью активность муравьи развивают на периферии территории (далее 10 метров от гнезда). Сбор строительного материала производится в пределах двухметровой полосы вдоль дорог.
Захаров справедливо отмечает сходство территориального поведения муравьев с таковым у птиц и хищных млекопитающих.
Инстинкт и научение в поведении насекомых.
Долгие годы господствовало мнение будто насекомые и другие счленистоногие являются существами, поведением которых руководит жесткий «слепой инстинкт». Это представление укоренилось преимущественно под влиянием работ выдающегося французского энтомолога Ж.-А.Фабра, который умел своими блестящими исследованиями убедительно показать, что даже сложнейшие действия насекомых не являются проявлением «разума», а выполняются на врожденной, инстинктивной основе. Одностороннее развитие положений Фабра и привело к указанной, неверной оценке поведения насекомых, к отрицанию не только разумности их поведения, но и к отрицанию, или хотя бы умалению роли накопления индивидуального опыта, научения в их жизни.
Как мы уже видели, формирование любой формы ви-дотипичного, наследственно «закодированного», т.е. инстинктивного, поведения в онтогенезе всегда сопряжено в той или иной степени с какими-либо элементами индивидуально приобретаемого поведения, научения. О строго фиксированном инстинктивном поведении в «чистом виде» не приходится говорить даже относительно низших животных.
В полной мере это относится и к насекомым, инстинктивное поведение которых также совершенствуется научением. В этом состоит основная роль научения в жизни насекомых. Можно, очевидно, считать, что научение стоит у насекомых и других членистоногих «на службе» у инстинктивного поведения. Как и у других животных, инстинктивные движения (врожденные двигательные координации) у них генетически строго фиксированы. Инстинктивные же действия, инстинктивное поведение являются и у насекомых в той или иной степени пластичными благодаря включению в них благоприобретаемых компонентов.
В естественных условиях способность к накоплению индивидуального опыта проявляется у насекомых в неодинаковой степени в разных функциональных сферах. Чаще всего она связана с ориентацией в пространстве и пищедобывательной деятельностью. Примером могут служить отмеченные выше опыты по обучению пчел за пищевое подкрепление ориентироваться по различным рисункам. Другой пример — муравьи, которые очень легко (всего за 12—15 опытов) научаются проходить даже сложный лабиринт, но, насколько известно, не научаются действиям, лежащим за пределами указанных функциональных сфер. Такая специфическая направленность (и одновременно ограниченность) способности к научению является характерной особенностью научения у представителей всего типа членистоногих.
Роль научения в поведении насекомых наглядно выступает и в «танцах» пчел — этих высших представителей членистоногих. Отстаивая взгляд, будто насекомые, в том числе пчелы, являются «стимульно связанными, рефлекторными животными», американские ученые В.Детьер и Э.Стеллар заявляют, например, что выполнению и интерпретации сложного танца пчелы не обучаются. Вместе с тем, как показали советские исследователи Н.Г.Лопатина, И.А.Никитина, Е.Г.Чеснокова и другие, процессы научения не только уточняют, но и модифицируют коммуникационные способности пчелы в онтогенезе и расширяют набор сигнальных средств.
Более того, как установили названные исследователи, биологическая значимость сигнальной деятельности медоносных пчел определяется стереотипом условных рефлексов, приобретаемых в онтогенезе по мере освоения пространства и при общении в семье. Оказалось, что интерпретация передаваемой в танце информации о расстоянии и направлении полета к источнику пищи возможна лишь в том случае, если пчела до этого научилась соотносить местонахождение корма с характером информации, содержащейся в танце сборщиц. Кроме того, тактильный компонент танца (вибрации брюшка) не имеет врожденного сигнального значения. Последнее приобретается в онтогенезе также условно-рефлекторным путем: пчелы, не имевшие в онтогенезе контактов (пищевых) с танцовщицей, не в состоянии интерпретировать этот существеннейший элемент танца. Следовательно, каждая пчела должна в основном научиться «понимать» язык танца. С другой стороны, образование временных связей оказалось важным и для формирования самой способности к выполнению танцев.
Таким образом, нет неизменных форм поведения даже там, где прежде всего требуется стереотипность, — в сигнальных позах и телодвижениях. Даже такое врожденное коммуникативное поведение, как «танцы» пчел, не только дополняется и обогащается процессами научения, не только переплетается с ними, но и формируется в комплексе с индивидуально приобретаемыми элементами поведения.
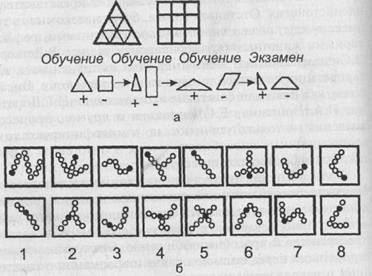
Рис. 41. Изучение способности медоносной пчелы к зрительному обобщению (опыты Мазохина-Поршнякова). Обозначения: а — общая схема проведения опытов; вверху — тестовые фигуры, внизу _ последовательность отдельных этапов формирования реакции на обобщенные признаки треугольника и четырехугольника (+ = пищевое подкрепление); б — опознавание рисунков по локальному признаку. В каждом опыте предлагалась на выбор одна пара из верхнего и нижнего ряда рисунков; подкреплялись только фигуры из верхнего ряда
Конечно, медоносная пчела занимает среди насекомых исключительное положение, и далеко не у всех представителей этого огромного класса психическое развитие достигает такой высоты. Об исключительных психических качествах медоносной пчелы свидетельствуют, в частности, экспериментальные данные, говорящие о наличии у нее аналогов некоторых психических функций высших позвоночных. Речь идет об установленной Мазохиным-Поршняковым высокоразвитой способности пчелы к зрительным обобщениям, например, типа «треугольник» и «четырехугольник» (независимо от конкретной формы, соотношения размеров и взаимной ориентации фигур) (рис. 41, а), «двуцветность» и др. В одной из серий опытов пчелам предлагалось выбрать из попарно предъявляемых фигур те, у которых один локальный признак (зачерченный кружочек) находился на конце цепочки из кружочков независимо от длины и формы этих цепей (рис. 41, б). Со всеми предложенными им задачами, даже в наиболее сложных вариантах, пчелы вполне справлялись. При этом отмечалась большая пластичность, нестандартность поведения, что экспериментатор справедливо связывает с непрерывной изменчивостью условий среды (непостоянство освещения, взаимного расположения, формы, окраски и многих других признаков компонентов среды), при которых этим насекомым приходится добывать пищу. Мазохин-Поршняков приходит к выводу, что выбор незнакомого объекта на основе обобщенных зрительных образов (иногда неправильно обозначаемых им как «понятия») есть свидетельство нестандартного использования пчелами индивидуального опыта, его применения в новой ситуации, отличной от обстановки первоначальной выработки соответствующего навыка.
Таким образом, здесь справедливо подчеркивается наличие и значение факта переноса определенного навыка в новую ситуацию и решение сложной задачи на основе индивидуального опыта, зафиксированного в виде обобщенного зрительного представления. В этом отношении мы действительно находим уже у пчел психические способности, аналогичные тем, которые относятся к предпосылкам интеллектуальных действий высших позвоночных животных. Однако одних этих предпосылок недостаточно для интеллектуального поведения, мышления животных, особенно если смотреть на эти высшие психические функции животных как на ступень по направлению к зарождению человеческого сознания. Поэтому описанные способности пчел не могут служить и критерием для признания у них мышления и уж во всяком случае не приходится говорить о наличии у пчелы рассудочной деятельности, хотя бы и в элементарной форме, как толкует результаты своих исследований Мазохин-Поршняков. Признавая у высших животных наличие своеобразных мыслительных способностей, интеллекта, нужно со всей определенностью отдавать себе отчет в том, что рассудок, т.е. разум, сознание как качественно иная категория психического отражения не присущи ни одному из животных, а только человеку.
Общая характеристика изшего уровня перцептивной психики.
На низшем уровне перцептивной психики уже представлены все те прогрессивные признаки, которые характеризуют перцептивную психику вообще, но во многих отношениях поведение относящихся сюда животных носит и примитивные черты, сближающие его с поведением нижестоящих животных. Так, основную роль играет ориентация поведения по-прежнему по отдельным свойствам предметов, но не по предметам как таковым: предметное восприятие явно играет еще подчиненную роль в общем поведении. Равным образом в последнем преобладают ригидные, «жестко запрограммированные» элементы поведения над гибкими, благоприобретаемыми и т.д.
С другой стороны, мы констатируем на этом уровне четко выраженный активный поиск положительных раздражителей, т.е. положительное таксисное поведение получает мощное развитие. Налицо все виды высших таксисов, включая мнемотаксисы. Последние играют в пространственной ориентации особенно существенную роль, и именно индивидуальном заучивании ориентиров проявляется в аибольшей степени способность к благоприобретаемому изменению видового поведения, к научению.
Вместе с тем важно подчеркнуть, что хотя у рассматриваемых здесь животных, в частности насекомых, накопление индивидуального опыта, научение играют существенную роль, наблюдается и определенная противоречивость в процессах научения, сочетания прогрессивных и примитивных черт. Специфическая направленность, приуроченность этих процессов к определенным функциональным сферам, как и само подчиненное положение, которое занимает научение по отношению к инстинктивному поведению, несомненно, указывают на переходное положение данного уровня психического развития между элементарной сенсорной и развитой перцептивной психикой.
Но, как мы видели, это никоим образом не означает, что насекомым, как и другим представителям рассматриваемой группы животных, недостает пластичности поведения. Наоборот, и здесь в полной мере проявляется общая закономерность, что усложнение инстинктивного поведения неизбежно сочетается с усложнением процессов научения (и наоборот). Только такое сочетание обеспечивает подлинный прогресс психической деятельности.
Инстинктивное поведение представлено на рассматриваемом уровне психического развития уже весьма развитыми новыми категориями: групповое поведение, общение, ритуализация. Особую сложность приобретают формы общения у видов, живущих огромными семьями, из которых лучше всего изучены пчелы. Язык пчел, этих высших представителей членистоногих, относится к наиболее сложным формам общения, которые вообще существуют в животном мире. Возможно, что новые исследования познакомят нас с поразительными психическими способностями и других насекомых, но на сегодняшний день пчелы представляются нам наиболее развитыми в этом отношении. Наиболее сложные формы инстинктивного поведения закономерно сочетаются у них с наиболее разнообразными и сложными проявлениями научения, что обеспечивает е только исключительную согласованность действий всех членов пчелиной семьи, но и максимальную пластичность поведения особи. Психические способности пчел (как и некоторых других высших насекомых) в некоторых отношениях, очевидно, уже выходят за рамки низшего уровня перцептивной психики.
В ином направлении, чем у членистоногих, шло развитие психической активности у головоногих моллюсков. По некоторым признакам они приблизились к ветви, ведущей к позвоночным, о чем уже свидетельствуют их крупные размеры и отмеченные выше особенности строения нервной системы и особенно зрительного рецептора, что непосредственно связано с резким увеличением скорости движения по сравнению с другими моллюсками.
Поведение головоногих еще совершенно недостаточно изучено, но уже известны многие примечательные их способности. Прежде всего они выделяются существенным усложнением инстинктивного поведения. У головоногих уже встречаются территориальное поведение (занятие и защита индивидуальных участков), «агрессивность», которая только намечается у высших червей, групповое поведение (стайная жизнь кальмаров и каракатиц), в сфере размножения появляются ритуализованные формы поведения, что находит свое воплощение в видоспецифическом «ухаживании» самцов за самками. Все это присуще только высшим животным, помимо головоногих, членистоногим и позвоночным.
Рядом исследователей обращалось особое внимание на весьма развитое у осьминогов «любопытство», что выражается в обследовании ими биологически «бесполезных» предметов, а также на их высокоразвитые манипуляцион-ные и конструктивные способности. Эти способности проявляются в строительстве валов и убежищ из камней, панцирей крабов, раковин устриц и т.п. Этот строительный материал осьминог подбирает, переносит й: укрепляет «руками». Иногда такие гнезда представляют собой закрытые со всех сторон строения. По некотбрым, правда еще. не подтвержденным, наблюдениям, осьминоги способны и к орудийным действиям, пользуясь камнями для защиты.
Очень важным представляется и то обстоятельство, что впервые у головоногих появляется способность к установлению контактов с человеком, к общению с ним, результатом чего является возможность подлинного приручения этих животных (в отличие от насекомых!).
Таким образом, головоногие, оставив далеко позади других моллюсков, как и вообще всех низших беспозвоночных, достигли, несомненно, высокого уровня психического развития, во многом сблизившись с позвоночными животными.
Вместе с тем и у головоногих наблюдается та же противоречивость в способности к научению, что и у насекомых. Так, например, у осьминога вообще хорошо развита способность к научению на зрительные и тактильные стимулы, но в ряде случаев он оказывается не в состоянии решить, казалось бы, несложные задачи. Особенно это относится к преодолению преград: осьминог неспособен находить обходной путь, если приманка (краб) располагается за прозрачной преградой (в стеклянном цилиндре или за проволочной сеткой). Тщетно пытаясь овладеть приманкой в прямом направлении, осьминог не хватает ее сверху, через край. Правда, по Бойтендайку, некоторые осьминоги все же способны решить несложные задачи обходного пути. При этом, очевидно, большое значение имеет прежний опыт особи. Другие головоногие уступают осьминогу по своим психическим способностям.
Конечно, при оценке подобных экспериментов необходимо иметь в виду, что здесь ставятся биологически неадекватные, а поэтому и неразрешимые задачи: в естественных условиях осьминог никогда не оказывается в ситуации, когда непосредственно зримая жертва оказывается недосягаемой. К тому же задачи обходного пути относятся к разряду весьма сложных — с ними не справляются не только черепахи, но и куры (в биологически несравненно более адекватных условиях). Тем не менее следует думать, что в психической деятельности головоногих действительно сочетаются прогрессивные черты, сближающие их с позвоночными, с примитивными — наследством низших моллюсков. К примитивным чертам относится и известный «негативизм» научения: головоногие легче научаются избегать неприятных раздражений, чем находить благоприятные. И в этом нетрудно усмотреть общность с поведением животных, обладающих элементарной сенсорной психикой.
На низшем уровне перцептивной психики находится, очевидно, хотя бы отчасти и ряд представителей низших позвоночных. Однако в корне различные строение и образ жизни членистоногих и позвоночных являются причиной того, что и их поведение и психика, в сущности, несопоставимы. Так, одной из отличительных особенностей насекомых являются их малые по сравнению с позвоночными размеры. В связи с этим окружающий насекомое мир представляет собой нечто совершенно особое: это не микромир простейших, но и не макромир позвоночных. Человеку трудно себе представить этот мир насекомых с его (с нашей точки зрения) микроландшафтами, микроклиматами и т.д. Хотя насекомые живут рядом и вместе с нами, они живут в совершенно иных условиях температуры, освещения и т.п. Уже поэтому психическое отражение действительности не может у насекомых не быть принципиально иным, чем у позвоночных, да и у большинства других беспозвоночных.
Но поскольку наиболее общие признаки психического отражения, характерные для данного его уровня, присущи всем упомянутым животным, можно, очевидно, по поводу рассмотренных нами в качестве примера насекомых сказать, что мы имеем здесь дело с типичными проявлениями низшего уровня перцептивной психики, но в формах, отвечающих тем особым условиям жизни этих животных, которые отмечались выше.
Высший уровень развития перцептивной психики.
В мире животных процесс эволюции привел к трем вершинам: позвоночные, насекомые и головоногие моллюски. Соответственно высокому уровню строения и жизнедеятельности этих животных мы наблюдаем у них и наиболее сложные формы поведения и психического отражения. Представители всех трех «вершин» способны к предметному восприятию, хотя, очевидно, только у позвоночных эта способность получила полное развитие. У остальных двух групп перцепция развивалась своеобразными путями и качественно отличается от таковой у позвоночных. Аналогично обстоит дело и с другими решающими критериями стадии перцептивной психики, не говоря уже о том, что высшего уровня перцептивной психики достигли в процессе эволюции вообще только представители позвоночных, и то явно не все. Только у высших позвоночных обнаруживаются все наиболее сложные проявления психической деятельности, которые вообще встречаются в мире животных.
Сравнивая беспозвоночных с позвоночными, необходимо также учесть, что ни головоногие, ни членистоногие не имеют никакого отношения к предкам позвоночных. Путь, ведущий к этим вершинам, отклонился от пути к третьей вершине еще на очень раннем этапе эволюции животного мира. Поэтому высокое развитие морфологических и поведенческих признаков, отмеченных нами у этих животных, является при сравнении с позвоночными лишь аналогией, обусловленной мощным повышением общего уровня жизнедеятельности, характерным для всех трех групп животных.
С филогенетической точки зрения для нас значительно больший интерес могли бы представить иглокожие, которые так же, как и позвоночные, относятся к вторич-норотым в отличие от первичноротых, у которых центральная нервная система располагается на брюшной стороне тела и к которым, в частности, относятся моллюски и членистоногие. Однако и современные иглокожие (морские ежи, звезды и т.д.) являются не предками позвоночных, а лишь боковой ветвью представленной весьма специализированными формами с примитивным поведением, характерным для элементарной сенсорной психики.
На этом уровне находятся и низшие хордовые, которые вместе с позвоночными (или «черепными») составляют тип хордовых. К низшим хордовым относятся оболочники и бесчерепные. Оболочники — морские животные, часть которых ведет неподвижную жизнь (асуи-дия). Бесчерепные представлены всего двумя семействами с тремя родами мелких морских животных, наиболее известное из которых — ланцетник. У бесчерепных и позвоночных общим признаком является внутренний осевой скелет, имеющий вид сплошного (хорда ланцетника) или членистого (позвоночник) стержня, над которым располагается центральная нервная система, имеющая форму трубки. Позвоночные подразделяются на классы кругло-ротых (миноги и миксины), рыб, земноводных, пресмыкающихся, птиц, млекопитающих. К высшим позвоночным относятся только последние два класса, в пределах которых, следовательно, и обнаруживаются проявления высших психических способностей животных. Что же касается низших позвоночных, то здесь намечаются промежуточные ступени психического развития, характеризующиеся разными сочетаниями элементов низшего и высшего уровней перцептивной психики.
У позвоночных нервная трубка образует в головном отделе вздутия, превращающиеся в ходе эмбриогенеза в головной мозг. Уже у наиболее примитивных позвоночных, у круглоротых, имеются все пять отделов головного мозга (продолговатый, задний, средний, промежуточный и передний мозг). Процесс дифференциации и прогрессивного развития мозговых структур досгигает, как известно, своей вершины у млекопитающих, причем не только в переднем мозгу (большие полушария и их кора), но и в стволовой части головного мозга, где формируются, в частности, центры высших форм инстинктивного поведения.
Что касается млекопитающих, то они вообще занимают особое место в эволюции животного мира, ибо, как указывал Северцов, «млекопитающие очень редко приспособляются к быстро наступающим переменам в окружающей среде (например, к новым врагам, к новой добыче и т.д.) обычным путем, то есть путем медленного изменения своих органов и их функций. Гораздо чаще это происходит путем быстрого изменения прежних привычек и навыков и образования новых, приспособленных к новым условиям среды. Здесь впервые выступает на сцену совершенно новый и необычайно важный фактор адаптивной эволюции позвоночных животных, а именно их психика»1.
Не нашли, что искали? Воспользуйтесь поиском: