ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Негистоновые белки хроматина 1 страница
В ядре эукариотической клетки присутствуют сотни самых разнообразных ДНК-связывающих негистоновых белков. Каждый белок комплементарен определённой последовательности нуклео-тидов ДНК (сайт ДНК). К этой группе относят семейство сайт-специфических белков типа "цинковые пальцы". Каждый "цинковый палец" узнаёт определённый сайт, состоящий из 5 нуклеотидных пар. Другое семейство сайт-специфических белков - гомодимеры. Фрагмент такого белка, контактирующий с ДНК, имеет структуру "спираль-поворот-спираль" (см. раздел 1). К группе структурных и регуляторных белков, которые постоянно ассоциированы с хроматином, относят белки высокой подвижности (HMG-белки - от англ, high mobility gel proteins). Они имеют молекулярную массу менее 30 кД и характеризуются высоким содержанием заряженных аминокислот. Благодаря небольшой молекулярной массе HMG-белки обладают высокой подвижностью при электрофорезе в полиакриламидном геле. К негистоновым белкам принадлежат также ферменты репликации, транскрипции и репарации. При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить нуклео-сом преобразуется в высококонденсированный комплекс белков и ДНК. Образованная структура в 10 000 раз короче исходной молекулы ДНК.
57) Дезоксирибонуклеи́новая кислота́ (ДНК) — один из двух типов нуклеиновых кислот, обеспечивающих хранение, передачу из поколения в поколение и реализацию генетическойпрограммы развития и функционирования живых организмов. Основная роль ДНК в клетках — долговременное хранение информации о структуре РНК и белков.В клетках эукариот (например, животных или растений) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
С химической точки зрения, ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков, нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы. В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула спирализована. В целом структура молекулы ДНК получила название «двойной спирали».
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином. Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.
58) Основные свойства генетического кода
· Код триплетен. В состав РНК входят 4 нуклеотида: А, Г, Ц, У. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 АМК зашифрована последовательностью трех нукл-ов, называемых триплетом или кодоном
· Код непрерывный. Это означает, что в коде отсутствуют сигналы, показывающие конец одного кодона и начало следующего. Поэтому рамка считывания должна быть правильно установлена в начале прочтения м-лы иРНК и затем двигаться последовательно от одного триплета к следующему. Если исходная рамка считывания оказалась «сбитой» в результате делеции или вставки на 1 или 2 нуклеотида, или же если рибосома случайно «пропустит» один нуклеотид в иРНК, все последующ кодоны выйдут из правильной рамки и это приводит к обр-нию белка с искаженной АМКной послед-стью.
· Код неперекрывающийся. В случае неперекрывающегося триплетного кода каждая группа из трех нукл-ов кодирует только одну АМКу
· Код вырожденный. Слово «вырожденность» – это матем термин, означающий в данном случае, что 1 АМКе может соотв больше одного кодона. Вырожденность кода вовсе не означает его несовершенство, поскольку нет ни одного кодона, к-рый бы кодировал несколько АМК. В физиологич усл код
· Однозначен: каждый кодон обозначает только одну АМКу. Вырожденность кода для различных АМК разная. Исключ сост метионин (AUG) и триптофан (UGG), кодирующиеся одним кодоном. Эти 2 АМКы встречаются в белках достаточно редко. Кодоны для гидрофобных АМК, например фенилаланина, лейцина, изолейцина и валина, различаются только одним основанием. Аналогичная ситуация наблюдается и для кодонов серина и треонина или аланина и глицина. При такой организации кода случайно возникшая замена основания с большей вероятностью, чем при случайном подборе кодонов, приведет к замене на сходную по свойствам АМКу или же замены не произойдет вовсе.
· Код универсальный. Ген код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушки и человека одни и те же триплеты кодируют одни и те же АМКы.
· Отсутствие знаков препинания
· Колинеарность – соответ послед-ти кодонов ДНК послед-ти АМК в белке.
· Бессмысленные кодоны: УАА, УАГ, УГА
59) мРНК, строение и роль
мРНК (синоним — информацио́нная РНК, иРНК) — РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков.мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.Длина типичной зрелой мРНК составляет от нескольких сотен до нескольких тысяч нуклеотидов. Эукариотические молекулы мРНК часто требуют сложной обработки и транспортировки из ядра — места синтеза мРНК, на рибосомы, где происходит трансляция, в то время как прокариотические молекулы мРНК этого не требуют и синтез РНК у них сопряжён с синтезом белка.Зрелая мРНК состоит из нескольких участков, различающихся по функциям: «5' кэп», 5' нетранслируемая область, кодирующая (транслируемая) область, 3' нетранслируемая область и 3' полиадениновый «хвост».5' кэп — это модифицированный гуанидиновый нуклеотид, который добавляется на 5' (передний) конец незрелой мРНК.Кодирующие области состоят из кодонов — следующих непосредственно друг за другом последовательностей из трёх нуклеотидов, каждая из которых соответствует в генетическом коде определённой аминокислоте или началу и концу синтеза белка.Нетранслируемые области — участки РНК, расположенные до старт-кодона и после стоп-кодона, которые не кодируют белок. Они называются 5'-нетранслируемая область и 3'-нетранслируемая область, соответственно.Длинная последовательность адениновых оснований, которая присутствует на 3' «хвосте» мРНК эукариот, синтезируется ферментом полиаденилат-полимеразой.Кроме первичной структуры (последовательности нуклеотидов), мРНК обладает вторичной структурой. Вторичная структура мРНК основана на внутримолекулярных взаимодействиях (линейная молекула «складывается», и водородные связи возникают между разными участками одной и той же молекулы).Осн назнач мРНК – переписывание ген инф-ции о первич стр-ре кодируемых белков с ДНК и участие в качестве матрицы в рибосомальном биосинтезе белков и пептидов.
60) тРНК, биороль
ТРАНСПОРТНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ- низкомолекулярные РНК, осуществляющие перенос аминокислотных остатков к матричной РНК (мРНК) при трансляции. Присутствуют в цитоплазме и митохондриях. Цитоплазматич. тРНК состоят из одной полирибонуклеотидной цепи, включающей 74-95 нуклеотидных остатков, митохондриальные тРНК немного короче. Для всех молекул тРНК характерно присутствие остатка фосфорной к-ты на 5'-конце, наличие последовательности ССА—ОН (С-остаток цитидина, А-аденозина) на 3'-конце и неск. консервативных нуклеотидных остатков, рассеянных вдоль цепи и занимающих во всех тРНК одни и те же положения.Отличит. особенность тРНК-присутствие в молекуле минорных нуклеозидов (миноров), Большинство миноров влияет на стабильность пространственной структуры и(или) на адапторную ф-цию тРНК-способность каждой тРНК узнавать свой кодон. Более половины пуриновых и пиримидиновых оснований тРНК с помощью водородных связей образуют внутрицепочечные пары по принципу комплементарности (A-U, G-C, G-U; U и G-соотв. остатки уридина и гуанозина), формируя 4 двухспиральных участка. Эти короткие спирали чередуются с участками неспаренных оснований, в результате чего нуклеотидная цепь образует 3 петли. Таким образом формируется вторичная структура, получившая назв. клеверного листа. В ней выделяют: акцепторную ветвь; дигидроуридиловую ветвь (шпильку); антикодоновую ветвь (шпильку); тимидилпсевдоуридиловую ветвь, или Ty-шпильку. Число генов, кодирующих тРНК для одной и той же аминокислоты, может различаться у разных организмов более чем на порядок. Помимо акцепторно-адапторной ф-ции в белковом синтезе, мн. тРНК выполняют роль затравки при обратной транскрипции (синтезе ДНК на РНК-матрице). На 3'-концах РНК мн. вирусов растений присутствуют тРНК-подобные структуры, обладающие акцепторной активностью. Нек-рые тРНК участвуют в биосинтезе пеп-тидогликанов (компонентов внеш. оболочки нек-рых бактерий), в переносе аминокислот через внеш. мембрану клеток, в регуляции биосинтеза ряда аминокислот, в посттрансляционной модификации белков (перенос аминокислотного остатка от аминоацил-тРНК на N-конец полипептидной цепи под действием ферментов аминоацил-тРНК-протеин трансфераз), а также во внутриклеточной деградации белков. Имеются данные об участии тРНК как кофактора в р-ции восстановления глутаминовой к-ты при биосинтезе хлорофилла.
61. Какое минимальное число нуклеотидных пар содержится в гене, кодирующем полипептид, состоящий из 124 аминокислотных остатков? Почему число нуклеотидных пар может оказаться гораздо больше, чем в ответе?
124*3+3*2=378. Число нуклеотидных пар может оказаться намного больше потому что есть некодирующие участки, такие как промотер, терминатор, интроны.
62. Как может повлиять изменение одного азотистого основания в мРНК на аминокислотную последовательность полипептида?
При изменении одного азотистого основания в мРНК аминокислотная последовательность полипептида также изменится, поскольку
63. Макроэргические соединения: АТФ и другие нуклеозидтрифосфаты, креатинфосфат, аргининфосфат и другие. Напишите структурную формулу АТФ. Укажите макроэргические связи.
Макроэргические молекулы (макроэрги) — биологические молекулы, высокоэнергетические природные соединения, которые способны накапливать и передавать энергию в ходе реакции.
Нуклеозидтрифосфаты (Nucleoside triphosphate, NTP) — это нуклеозиды с тремя фосфатами. Природные нуклеозидтрифосфаты представлены аденозинтрифосфатом (ATP), гуанозинтрифосфатом (GTP), цитидинтрифосфатом (CTP), тимидинтрифосфатом (TTP) и уридинтрифосфатом (UTP). Данные термины означают, что нуклеотиды содержат сахар рибозу.
Нуклеотиды, содержащие сахар дезоксирибозу, имеют приставку дезокси- в имени и d- в сокращении: дезоксиаденозинтрифосфатом (dATP), дезоксигуанозинтрифосфатом (dGTP), дезоксицитидинтрифосфатом (dCTP), дезокситимидинтрифосфатом (dTTP) и дезоксиуридинтрифосфатом (dUTP).
Важные представители фосфогенов - креатинфосфат (у большинства позвоночных) и аргининфосфат (у многих беспозвоночных) - являются производными гуанидина. В их молекулах атомы фосфора непосредственно связаны с азотом. Большое значение стандартной свободной энергии их гидролиза обусловлено трудностями стабилизации гуанидиновой и фосфатной групп в молекулах. В результате гидролиза отщепляется фосфатная группа, а продукты реакции стабилизируются. Фосфогены выполняют роль аккумуляторов энергии и возникают при фосфорилировании креатина и аргинина с помощью АТФ. В случае необходимости они могут освобождать энергию и ресинтезировать АТФ из АДФ. Креатинфосфат — продукт обратимого метаболического N-фосфорилирования креатина, являющийся, подобно АТФ, высокоэнергетическим соединением. Однако, в отличие от АТФ, гидролизуемой по пирофосфатной связи O-P, креатин гидролизуется по фосфамидной связи N-P, что обуславливает значительно больший энергетический эффект реакции. Так, при гидролизе изменение свободной энергии для креатина ~ −43 кДж/моль, в то время как при гидролизе АТФ до АДФ ~ −30 кДж/моль. Креатинфосфат содержится преимущественно в возбудимых тканях (мышечная и нервная ткани) и его биологической функцией является поддержание постоянной концентрации АТФ за счёт обратимой реакции перефосфорилирования:
креатинфосфат + АДФ ⇔ креатин + АТФ
Структурная формула АТФ (красным обозначены макроэрг. связи)

64. Какие химические соединения называются макроэргическими? Напишите формулы нуклеотидов, являющихся макроэргическими соединениями. Какова их биологическая роль.
Макроэргические молекулы (макроэрги) — биологические молекулы, высокоэнергетические природные соединения, которые способны накапливать и передавать энергию в ходе реакции.
Аденинтрифосфорная к-та.
Источник энергии:
1) для реакций ассимиляции (синтеза веществ)
2) для активного транспорта веществ
3) для движения
4) для свечения (ночесветки и пр.)
5) для работы элетрических органов (например, у электрических угрей и скатов).
Кроме того является источником фосфатных группировок для синтеза различных содержащих эту группировку веществ.
АТФ не только источник энергии, а т.ж.:
1) Он учавствует в синтезе нуклеиновых кислот,
2) Учавствует в передаче гормонального сигнала в клетку,
3) Является ингибитором некоторых ферментов
4) Медиатором в некоторых рецепторах.

Кроме АТФ существуют и другие макроэргические соединения на основе нуклеотидов: ГТФ или гуанозинтрифосфат (содержит гуанин; участвует в биосинтезе белков, глюкозы),

УДФ (содержит урацил; участвует в синтезе полисахаридов).

Цитидинтрифосфат (Она используется как источник энергии и как кофермент в метаболических реакциях, таких как синтез глицерофосфолипидов и гликозилирование белков. Цитидинтрифосфат участвует в синтезе практически всех фосфолипидов и является источником цитидина в процессе синтеза рибонуклеиновой кислоты, так же как дезоксицитидинтрифосфат является источником дезоксицитидина в синтезе ДНК. Цитидинтрифосфат участвует в процессе образования АТФ в качестве донора фосфатных остатков для АДФ.)

Нуклеотиды способны образовывать циклические формы, например, цАМФ, цЦМФ, цГМФ. Циклические нуклеотиды выполняют роль регуляторов различных физиологических процессов.
65. Классификация и номенклатура углеводов. Особенности строения моносахаридов.
Углеводы делятся на:
А) Моносахариды:
- Альдозы;
- Кетозы.
Б) Олигосахариды (дисахариды, трисахариды и т.д.)
В) Полисахариды:
- Гомополисахариды;
- Гетерополисахариды.
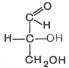
Моносахариды можно рассматривать как производные многоатомных спиртов, содержащие карбонильную (альдегидную или кетонную) группу. Если карбонильная группа находится в конце цепи, то моносахарид представляет собой альдегид и называется альдозой; при любом другом положении этой группы моносахарид является кетоном и называется кетозой. Простейшие представители моносахаридов – триозы: глицеральдегид и диоксиацетон. При окислении первичной спиртовой группы трехатомного спирта – глицерола – образуется глицеральдегид (альдоза), а окисление вторичной спиртовой группы приводит к образованию диоксиацетона (кетоза).

Стереоизомерия моносахаридов. Все моносахариды содержат асимметричные атомы углерода: альдотриозы – один центр асимметрии, альдотетрозы – 2, альдопентозы – 3, альдогексозы – 4 и т.д. Кетозы содержат на один асимметричный атом меньше, чем альдозы с тем же числом углеродных атомов. Следовательно, кетотриоза диоксиацетон не содержит асимметричных атомов углерода. Все остальные моносахариды могут существовать в виде различных стереоизомеров. Общее число стереоизомеров для любого моносахарида выражается формулой N = 2n, где N – число стереоизомеров, а n – число асимметричных атомов углерода. Как отмечалось, глицеральдегид содержит только один асимметричный атом углерода и поэтому может существовать в виде двух различных стереоизомеров. Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у асимметричного атома углерода расположена с правой стороны, принято считать D-глицеральдегидом, а зеркальное отражение – L-глицеральдегидом:

D-глицеральдегид L-глицеральдегид
Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде 16 стереоизомеров (24), представителем которых является, например, глюкоза. Для альдопентоз и альдотетроз число стереоизомеров равно соответственно 23 = 8 и 22 = 4. Все изомеры моносахаридов подразделяются на D- и L-формы (D- и L-конфигурация) по сходству расположения групп атомов у последнего центра асимметрии с расположением групп у D- и L-глицеральдегида. Природные гексозы: глюкоза, фруктоза, манноза и галактоза – принадлежат, как правило, по стереохимической конфигурации к соединениям D-ряда. Известно, что природные моносахариды обладают оптической активностью.
Циклические (полуацетальные) формы моносахаридов. Любой моносахарид с конкретными физическими свойствами (температура плавления, растворимость и т.д.) характеризуется специфической величиной удельного вращения  . Установлено, что величина удельного вращения при растворении любого моносахарида постепенно меняется и лишь при длительном стоянии раствора достигает вполне определенного значения. Например, для свежеприготовлен-ного раствора глюкозы = +112,2°, после длительного стояния раствора эта величина достигает равновесного значения = +52,5°. Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией. Очевидно, мутаротация должна вызываться изменением асимметрии молекулы, а следовательно, трансформацией ее структуры в растворе. Явление мутаротации имеет объяснение. Известно, что альдегиды и кетоны легко и обратимо реагируют с эквимолярным количеством спирта с образованием полуацеталей:
. Установлено, что величина удельного вращения при растворении любого моносахарида постепенно меняется и лишь при длительном стоянии раствора достигает вполне определенного значения. Например, для свежеприготовлен-ного раствора глюкозы = +112,2°, после длительного стояния раствора эта величина достигает равновесного значения = +52,5°. Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией. Очевидно, мутаротация должна вызываться изменением асимметрии молекулы, а следовательно, трансформацией ее структуры в растворе. Явление мутаротации имеет объяснение. Известно, что альдегиды и кетоны легко и обратимо реагируют с эквимолярным количеством спирта с образованием полуацеталей:

Полуацеталь
Реакция образования полуацеталя возможна и в пределах одной молекулы, если это не связано с пространственными ограничениями. По теории А. Байера, внутримолекулярное взаимодействие спиртовой и карбонильной групп наиболее благоприятно, если оно приводит к образованию пяти- или шестичленных циклов. При образовании полуацеталей возникает новый асимметрический центр (для D-глюкозы это С-1). Шестичленные кольца сахаров называют пиранозами, а пятичленные – фуранозами. α-Форма – это форма, у которой расположение полуацетального гидроксила такое же, как у асимметричного углеродного атома, определяющего принадлежность к D- или L-ряду. Иными словами, в формулах с α-модификацией моносахаридов D-ряда полуацетальный гидроксил пишут справа, а в формулах представителей L-ряда – слева. При написании β-формы поступают наоборот. Таким образом, явление мутаротации связано с тем, что каждый твердый препарат углеводов представляет собой какую-либо одну циклическую (полуацетальную) форму, но при растворении и стоянии растворов эта форма через альдегидную превращается в другие таутомерные циклические формы до достижения состояния равновесия. При этом значение удельного вращения, характерное для исходной циклической формы, посте-пенно меняется. Наконец, устанавливается постоянное удельное вращение, которое характерно для равновесной смеси таутомеров.
66. Дайте определения следующим понятиям: оптические антиподы, эпимеры, диастереоизомеры, рацемическая смесь.
Напишите структурные формулы следующих моносахаридов:
эпимер D-глюкозы по С2;
аномер а,Б-глюкопиранозы.
Назовите эти моносахариды.
Антиподы оптические, пространственные изомеры с одинаковыми химическими и физическими свойствами, кроме направления (знака) вращения плоскости поляризации света. По структуре молекулы А. о. относятся друг к другу, как предмет к своему зеркальному изображению (см. Изомерия). Антипод, вращающий плоскость поляризации света вправо, называется право-вращающим [(+)-антиподом], а вращающий влево — левовращающим [(—)-антиподом]. Примером А. о. может служить глицериновый альдегид

Эпимеры - стереоизомеры, содержащие несколько хиральных центров, но различающиеся по конфигурации только одного из центров.
Диастереоизомеры - пространственные изомеры с несколькими асимметрическими центрами, у которых конфигурация части асимметрических атомов совпадает, а части - противоположна. В отличие от антиподов оптических, у которых конфигурация всех асимметрических центров соответственно противоположна и которые отличаются друг от друга только знаком оптического вращения при полном совпадении всех др. свойств, Д. отличаются не только величиной оптического вращения, но и другими физическими свойствами, например температурами плавления, растворимостью. Они отличаются также некоторыми химическими свойствами (скоростями реакций).
Рацемическая смесь - смесь в равных пропорциях двух энантиомеров в-в, проявляющих оптическую активность. Один из них - правовращающий, т.е. поворачивающий плоскость поляризации света, проходящего через него, в правую сторону; др. - левовращающий, отличается только направлением поворота. В рез-те оба эффекта взаимно гасят друг друга, и Р.с. не обнаруживает оптич. активности. Назв. происходит от рацемической винной к-ты, представляющей собой смесь равных кол-в D-(правовращающей) и L-(левовращающей) винных к-т.
эпимер D-глюкозы по С2 (D-манноза):

аномер а,Б-глюкопиранозы (на рисунке слева вверху)

67. Что такое мутаротация? Как можно объяснить преобладание [3- D-глюкопиранозы в растворе после стояния? Какие еще формы глюкозы находятся в растворе? Напишите формулы.
Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией.
Явление мутаротации имеет объяснение. Известно, что альдегиды и кетоны легко и обратимо реагируют с эквимолярным количеством спирта с образованием полуацеталей:
Полуацеталь
Реакция образования полуацеталя возможна и в пределах одной молекулы, если это не связано с пространственными ограничениями. По теории А. Байера, внутримолекулярное взаимодействие спиртовой и карбонильной групп наиболее благоприятно, если оно приводит к образованию пяти- или шестичленных циклов. При образовании полуацеталей возникает новый асимметрический центр (для D-глюкозы это С-1). Шестичленные кольца сахаров называют пиранозами, а пятичленные – фуранозами. α-Форма – это форма, у которой расположение полуацетального гидроксила такое же, как у асимметричного углеродного атома, определяющего принадлежность к D- или L-ряду. Иными словами, в формулах с α-модификацией моносахаридов D-ряда полуацетальный гидроксил пишут справа, а в формулах представителей L-ряда – слева. При написании β-формы поступают наоборот. Таким образом, явление мутаротации связано с тем, что каждый твердый препарат углеводов представляет собой какую-либо одну циклическую (полуацетальную) форму, но при растворении и стоянии растворов эта форма через альдегидную превращается в другие таутомерные циклические формы до достижения состояния равновесия. При этом значение удельного вращения, характерное для исходной циклической формы, посте-пенно меняется. Наконец, устанавливается постоянное удельное вращение, которое характерно для равновесной смеси таутомеров.
Например, известно, что в водных растворах глюкоза находится главным образом в виде α- и β-глюкопираноз, в меньшей степени – в виде α- и β-глюкофураноз и совсем небольшое количество глюкозы – в виде альдегидной формы. Следует подчеркнуть, что из различных таутомерных форм глюкозы в свободном состоянии известны лишь α- и β-пиранозы. Существование малых количеств фураноз и альдегидной формы в растворах доказано, но в свободном состоянии они не могут быть выделены вследствие своей неустойчивости.
В растворе глюкозы находятся следующие формы глюкозы:

68. Основные таутомерные формы глюкозы в растворе. Напишите их формулы. Что такое мутаротация?
Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией.

69. Какая функциональная группа глюкозы проявляет восстанавливающие свойства? Напишите схему реакции окисления глюкозы гидроксидом меди (II).
 +2Cu(OH)2->
+2Cu(OH)2->  +Cu2O+2H2O
+Cu2O+2H2O
Окисляется альдегидная группа (т.е. проявляет восст. свойства), выпадает красный осадок оксида меди (I), образуется глюконовая кислота. Реакция хорошо идет при слабом нагревании, при низкой температуре образуется синий комплекс без окисления альдегидной группы, поскольку глюкоза еще и многоатомный спирт.
70. Химические свойства моносахаридов (реакции с участием карбонильной и спиртовой групп, гликозидного гидроксила).
Реакции полуацетального гидроксила. Уже отмечалось, что моносахариды как в кристаллическом состоянии, так и в растворе в основном существуют в полуацетальных формах. Полуацетальный гидроксил отличается большей реакционной способностью и может замещаться другими группировками в реакциях со спиртами, карбоновыми кислотами, фенолами и т.д.
Продукт реакции называют гликозидом. Соответственно α- и β-изомерам моносахаридов существуют α- и β-глюкозиды. Например, при реакции метилового спирта с глюкозой (допустим, в β-пиранозной форме) в присутствии неорганических кислот образуется продукт алкилирования метил-β-D-глюкопиранозид:
СH3OH H2O

b-D-глюкопираноза Метил-b-D-глюкопиранозид
При действии на β-D-глюкопиранозу уксусной кислотой образуется продукт ацилирования ацетил-β-D-глюкопиранозид (по той же схеме).
Ацилированию и метилированию способны подвергаться и остальные группы моносахаридов, но при намного более жестких условиях. Если в реакцию вступают спирты, фенолы или карбоновые кислоты, продукты реакции называют О-гликозидами. Следовательно, метил-β-D-глюкопиранозид и ацетил-β-D-глюкопиранозид являются О-гликозидами (связь осуществляется через кислород). Природные О-гликозиды, большинство из которых образуется в результате жизнедеятельности растений, существуют преимущественно в β-форме.
Не нашли, что искали? Воспользуйтесь поиском: