ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Гликолиз. Р-ции. Регуляция. 1 страница
Реакции:
1)2ПВК+2АТФ+2СО2->(пируваткарбоксилаза)2 оксалоацетат+2фосфат неорганич.+2 АТФ
2)2 ацелоацетат+ 2 НАДН(Н+)->(малатдегидрогеназа) 2 малат+2НАД+
3)2 малат +2НАД+->(малатдегидрогеназа)2 ацелоацетат+2НАДН(Н+)
4)2 оксалоацетат+2 ГТФ->(фосфоенол-пируват-карбоксикиназа)2 фосфоенол-пируват+2ГТФ
5)2 фосфоенол-пируват->(енолаза)2 фосфоглицерат
6)2 фосфоглицерат->(фосфоглицератмутаза) 2 3-фосфоглицерат
7)2 3-фосфоглицерат +2АТФ->(фосфоглицераткиназа)2 1,3-дифосфоглицерат +2 АДФ
8)2 1,3-дифосфоглицерат +2 НАДН(Н+)->(глицеральдегид-3-фосфат-дегидрогеназа) 2 глицеральдегид-3-фосфат+2 фосфат нерганич.
9)Глицеральдегид-3-фосфат->(триозофосфат-изомераза) дигидрокси ацетон фосфат
10)Глицеральдегид-3-фосфат+дигидрокси-ацетон-фосфат->(фруктозодифосфат-альдалаза) фруктозо-1,6-дифосфат
11)Фруктозо-1,6-дифосфат+Н2О->(фруктозодифосфатаза) фруктозо-6-фосфат+фосфат неорганич.
12)Фруктозо-6-фосфат->(фосфоглико-изомераза) глюкозо-6-фосфат
13)Глюкозо-6-фосфат + Н2О->(глюкозо-6-фосфатаза) глюкоза+ фосфат неоганич.
Регуляция гликолиза: гликолиз стимулир-ся инсулином, повышающим кол-во молекул гексокиназы, фосфофруктокиназы, пируваткиназы. В печени активность глюкокиназы регулир-ся гормонами: активацию вызывает инсулин и андрогены, подавляют ее активность глюкокортикоиды и эстрогены.
57. Пентозофосфатный путь (ПФП) -это альтернативный путь окисл-я гл-зы в печени, мол.ж-зе и жир.тк. Фер-ты этого пути распол-ны в цит-ме. Сост.из 2 этапов: окисл-го и неокисл-го. В завис-ти от необх-ти он может осущ-ся по циклич-му или шунтирующему пути. Один из вариантов стехиометрии предпол., что из 3 мол-л гл-зо-6-фосфатов образ-ся 3 мол-лыCO2 и 3 мол-лы пяти углеродн.сахаров. Пентоза превр-ся 2 мол-лы глюкозо-6-фосфата и 1 мол-лу глицеральдегид фосфат. Акцептором протонов и электронов служит НАД. Р-ции: 1) Гл-зо-6-фосфат при участии фермента гл-зо-6-фосфат дегидрогеназа в присутствии НАДФ и кофакторовMn2+или Ca2+ обратимо превр-ся в 6-глюконолактон. Из него образ-ся 6-фосфоглюконат (фермент глюконолактонгидролаза). Кофакторы те же. 2) 6-фосфоглюконат окисл-ся 6 фосфоглюконатдегидрогеназой кофермент НАДФ, кофакторыMn2+, Ca2+. В обратим.р-ции до 3-кето-6-фосфоглюконата, при декарбоксилировании которого обр-ся рибулоза-5-фосфат. 3) Фермент рибулозо-5-фосфат эпимераза изм-т конфигурации мол-лы в обл.3 углер.атома превращая рибулозо-5 фосфат в ксилулозо-5-фосфат (обрат.р-ция). Альтер-но фермент рибозо-5-фосфат кетоизомераза обратимо превращает в рибулозо-5-фосфат в рибозо-5-фосфат. 4) Транскетолаза при помощи тиаминпирофосфата и Mg2+переносят 1 и 2 атомы углерода. Ксилозо-5-фосфат на рибозо-5-фосфат, в рез-те чего образ-ся седогетулозо-7-фосфат и глицеральдегид-6-фосфат. 5) Трансальдолаза катализирует перенос 3-х углер.части седогептулозо-7-фосфат на глицеральдегид-3-фосфат с образованием фруктозо-6-фосфата и эритрозо-4фосфата. 6) Транскетолаза при помощи тиамин пирофосфата и Mg2+ переносит 1-й и 2-ой атомы углерода ксилулозо-5-фосфата на эритрозо-4-фосфат с образованием фр-зо-6-фосфата и глицеральдегид-3- фосфата. Значение ПФП: 1. Он является главным источником восстановит-го экв-та НАДФН(Н+),который исполь-ся в биосинтезах,а также в реакциях нейтромирации активн.форм кислорода. 2. В ходе р-ций обр-ся рибоза-5-фосфат,который исп-ся для синтеза нукл к-т и носителей Е.(нуклеотидов). 3. В неокислит.части обр-ся сложн.сахара используемые при образование компонента межклеточного матрикса, углеводных компонентов рецептора и тд.
58. Биосинтез углеводов в тканях. Реакции глюконеогенеза и гликогеногенеза, углеводные и неуглеводные источники для глюконеогенеза, взаимоотношение процессов синтеза и распада гликогена. Биосинтез углеводов в тканях происходит аэробным (с доступом кислорода) и анаэробным (без доступа кислорода) путями. Анаэробный путь- гликолиз и гликогенолиз. Гликолиз - процесс распада глюкозы, протекающий без потребления О2.Конечный продукт- молочная к-та. В процесс гли-за образуется АТР. Суммарное ур-ие гли-за:C6H12+2АДФ+2Фн ->2CH3CH(OH)COOH+2H2O. Благодаря гликолизу организм человека опред-ый период может осуществлять ряд физиологических ф-ий в условиях недостаточности О2. Его ферменты способны связываться с мышечными белками. Гликогенолиз - расщепление начинается с гликогена. Начальная реакция - фосфорилиз гликогена- расщепление с присоединением фосфор. к-ты. В печени и почках имеется фермент- глюкоза-6- фосфотаза. Образовавшаяся глюкоза выходит из клетки в кровь. Это заключит. Стадия гликогенолиза в печени, приводящего к повышению содержания глюкозы в крови. Аэробный путь - окисление глюкозы до СО2 и Н2О, пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, то пируват превращается в лактат.Аэробный распад глюкозы можно выразить суммарным уравнением: С6Н12О6 + 6 О2 → 6 СО2 + Н2О + 2820 кДж/моль. Глюконеогенез -это образование глюкозы из таких неуглеводных предшественников лактата, циклокарбоновых к-т, глицерола, пропиата. Состоит из 10 р-ций- необратимые: 1,4,11,13, остальные обратимые.
1)2ПВК+2АТФ+2СО2->(пируваткарбоксилаза)2 оксалоацетат+2фосфат неорганич.+2 АТФ
2)2 ацелоацетат+ 2 НАДН(Н+)->(малатдегидрогеназа) 2 малат+2НАД+
3)2 малат +2НАД+->(малатдегидрогеназа)2 ацелоацетат+2НАДН(Н+)
4)2 оксалоацетат+2 ГТФ->(фосфоенол-пируват-карбоксикиназа)2 фосфоенол-пируват+2ГТФ
5)2 фосфоенол-пируват->(енолаза)2 фосфоглицерат
6)2 фосфоглицерат->(фосфоглицератмутаза) 2 3-фосфоглицерат
7)2 3-фосфоглицерат +2АТФ->(фосфоглицераткиназа)2 1,3-дифосфоглицерат +2 АДФ
8)2 1,3-дифосфоглицерат +2 НАДН(Н+)->(глицеральдегид-3-фосфат-дегидрогеназа) 2 глицеральдегид-3-фосфат+2 фосфат нерганич.
9)Глицеральдегид-3-фосфат->(триозофосфат-изомераза) дигидрокси ацетон фосфат
10)Глицеральдегид-3-фосфат+дигидрокси-ацетон-фосфат->(фруктозодифосфат-альдалаза) фруктозо-1,6-дифосфат
11)Фруктозо-1,6-дифосфат+Н2О->(фруктозодифосфатаза) фруктозо-6-фосфат+фосфат неорганич.
12)Фруктозо-6-фосфат->(фосфоглико-изомераза) глюкозо-6-фосфат
13)Глюкозо-6-фосфат + Н2О->(глюкозо-6-фосфатаза) глюкоза+ фосфат неоганич.
59. Биосинтез и распад гликогена. Регуляция обмена гликогена. Гликоген - разветвлённый полимер глюкозы. Наибольшая его концентрация обнаружена в печени и мышцах. Поскольку мышечная масса организма человека велика, то большая часть гликогена организма содержится в мышцах. Глюкоза из крови легко проникает в клетки органов и тканей, проходя ч/з биологические мембраны клеток. Как только глюкоза поступает в клетку, она метаболизируется в ней в результате первой химической реакции. Фосфорилирование глюкозы происходит в присутствии АТФ и фермента - гексокиназы. Глюкоза превращается в глюкозо-6-фосфат. Этот эфир глюкозы теперь будет использоваться в анаболических и катаболических реакциях. Глюкоза из клетки может выйти только после реакции гидролиза при участии глюкозо-6-фосфатазы. Этот фермент есть в печени, почках, эпителии кишечника. В других органах, тканях его нет. Процесс биосинтеза гликогена протекает в 4 стадии. Ферменты: 1) фосфорилаза, 2) фосфоглюкомутаза, 3) глюкоза-6-фосфатаза, 4) глюкокиназа, 5) глюкозо-1-фосфатуридил трансфелаза, 6) гликоген-синтетаза. Основная часть гликогена нах-ся в печени и истощается за сутки. Фермент гликогенсинтетаза катал-ся присоединением глюкозы. И сущ-ет в 2 формах: А) активная (дефосфолирована).Б) неактивная (фосфолирована). Фосфолирование катал-ся протеинкиназами, там присоединяется АМФ, синтез которой стим-ся адреналином и глюкагоном. Переход из активной формы в неактиную актив-ся гидрокартизоном, инсулином, глюкозой и ИНГИБИРУЕТСЯ гликогеном. Глюкоза подвергается фосфолированию при участии фермента гексокиназы, а в печени- глюкокиназы. глюкозо-6-фосфат->(фосфоглюкомутазыа) глюкозо-1-фос-фат. Сущ-ет 2 стадии- 1) Стадия синтеза глюкозо-1-фосфата: глюкозо-1-фосфат+УТФ (уридинтрифосфат)-> (УДФ-глюкоза) + пирофосфат. 2) стадия образования гликогена. C6H12O6->(гексокиназа)глюкоза-6-фосфат->(фосфомутаза)глюкоза-1-фосфат. Распад - это процесс расщепления гликогена. Сущ-ет 2 пути распада: 1) Гидролитический- осуществ-ся под действием фермента амилазы в печени в присутствии воды и выделяется свободная глюкоза. 2) Фосфоролитический- катализ-ся ферментом фосфорилазой. Это расщепление энергетически выгодно, т.к освободжается фосфолиривонная глюкоза. Главные ферменты, контролирующие метаболизм гликогена – гликогенфосфорилаза и гликогенсинтаза.
60. Глюконеогенез Регуляция. Реакции и роль в обмене вещ-в. Глюконеогенез -это образование глюкозы из таких неуглеводных предшественников,как лактата,промеж.продукты циклокарбон. К-т,глицерола,пропиата.
Реакции: Необратимые: 1,4,11,13!!!!
1)2ПВК+2АТФ+2СО2->(пируваткарбоксилаза)2 оксалоацетат+2фосфат неорганич.+2 АДФ
2)2 ацелоацетат+ 2 НАДН(Н+)->(малатдегидрогеназа) 2 малат+2НАД+
3)2 малат +2НАД+->(малатдегидрогеназа)2 ацелоацетат+2НАДН(Н+)
4)2 оксалоацетат+2 ГТФ->(фосфоенол-пируват-карбоксикиназа)2 фосфоенол-пируват+2ГДФ
5)2 фосфоенол-пируват->(енолаза)2 фосфоглицерат
6)2 фосфоглицерат->(фосфоглицератмутаза) 2 3-фосфоглицерат
7)2 3-фосфоглицерат +2АТФ->(фосфоглицераткиназа)2 1,3-дифосфоглицерат +2 АДФ
8)2 1,3-дифосфоглицерат +2 НАДН(Н+)->(глицеральдегид-3-фосфат-дегидрогеназа) 2 глицеральдегид-3-фосфат+2 фосфат нерганич.
9)Глицеральдегид-3-фосфат->(триозофосфат-изомераза) дигидрокси ацетон фосфат
10)Глицеральдегид-3-фосфат+дигидрокси-ацетон-фосфат->(фруктозодифосфат-альдалаза) фруктозо-1,6-дифосфат
11)Фруктозо-1,6-дифосфат+Н2О->(фруктозодифосфатаза) фруктозо-6-фосфат+фосфат неорганич.
12)Фруктозо-6-фосфат->(фосфоглико-изомераза) глюкозо-6-фосфат
13)Глюкозо-6-фосфат + Н2О->(глюкозо-6-фосфатаза) глюкоза+ фосфат неорганич.
Регуляция глюконеогенеза: Превращение метаболитов в глюконеогенезе связаны так,что один процесс активен,а второй неактивен. А вообще термодинамически выгодна их одновременная активация. 1.Ацетил-КоАактивирует,а АДФ ингибирует пируваткарбоксилазу(первый фермент глюконеогенеза). Когда в клетке накапливается митохондриальный ацетил-КоА,биосинтез глюкозы из пируватаусиливается.,одновременно подавляются окислительные декарбоксилирования пирувата. Скорость глюконеогенеза зависит от концентрации лактата и других предшественников глюкозы 2. Фруктоза-1,6-бисфосфотаза ингибируется АМФ. В тоже время АМФ является аллостерическим
активатором фосфофруктокиназы. При низкой концентрации АМФ и высокой концентрации АТФ происходит стимуляция глюконеогенеза. А когда величина АМФ низкая,в клетке наблюдается расщепление глюкозы. Фруктозо-2,6-бисфосфатаза активирует фосфо-фруктокиназу и ингибирует фруктозодифосфатазу
Роль глюконеогенеза в обмене углеводов: глюконеогенез не только обеспечивает органы и ткани глюкозой, но еще и перерабатывает образующийся в тканях лактат, препятствуя тем самым развитию лактат-ацидоза.
61. Нарушение обмена углеводов. Выделяют след.основныенарушения обмена угл-ов:1.гипер-и гипогликимия.2.болезни накопления гликогена. 3.гликозурия. 4.галактозимия. 5.лактозурия.6.мальтозурия. 7.фруктозурия-причиной может быть:а)фруктозная нагрузка;б)дефицит фруктокиназы (неспособность потреблять фруктозу);в)недостаток или частич. Неактивность фруктозодифосфатальдолазы в печени;следствие накопление фруктозо-1-фофата блокирующего потребление глюкозы.
Норм.конц.глюкозы в крови 3,3-6 ммоль/л. Гипергликимия может быть вызвана избыт.кол-ом глюкозы в пищи или сах.диабетом.Сах.диабет бывает первичн. или вторичный. 1.Первичный (идиопатический) подраздел.на: а)потенциальный(фамильная предрасположенность); б)латентный (скрытый)-проявл.при лечении картизоном, острых инфекциях, беременности;характерен диабетический тип кривой сахарной нагрузки; в)асимптоматический (химич) г)клинический-делится на:- первого типа -инсулинзависимый (ювенильный)-проявл.в первые 40 лет жизни и хар-ся резким снижением секреции инсулина вследствие атрофии бета-клеток.- второго типа- инсулин-независимый. Поражает людей тарше 40 лет,хар-сянекот.снижениемсекреции инсулина. 2.Вторичный. Причины: а)поврежд.поджел. железа, прихронич. панкреатите,опухолях и др.очевидныхзаболеваниях; б)присутствие антаганистовинсулина: избыточная секреция гормона роста или глюкокартикоидов; в)ингибирование секреции инсулина:секреция инсулина бета-клетками может подавляться при избыт.продуктахнорадреналина,что приводит к мобилизации гликогена в печени.Адреналин и норадреналин блокируют секрецию инсулина.Др.причины диабелта:1.частичное нарушение норм.структуры инсулина;2.повыш.уровень секреции инсулиназы; 3.частичн. наруш.норм. структуры инсулиновых рецепторов. Гипогликимия делится на: а)гипогликимияголодание. Причины: инсулинсекретирующие опухоли (поджел.железа), недостаточн.ф-ии гипофиза и надпочечников,болезни печени и болезни накопления гликогена.б)реактивная гипогликимия. Причины: передоз. инсулина, физич.нагрузка,прием лекарственных препаратов, приемалкоголя, частичная реакция желудка. Такое состояние может вызвать сульфанилмочевина. Глюкозурия делится на: 1.алиментарная(избыток глюкозы в пище);2.эмарганальная(стресс). Др.причины. 1.действие хим. препаратов(морфин,хлороформ); 2.диабет;3.низкий порог ренальной реабсорбции;4.остр.инфекции,сотряс.мозга.
62. Гликогенозы, причины, сущность, проявления заболевания. Значение нарушений активности глюкозо-6-фосфотазы, кислой альфа-глюкозидазы, фосфорилазы, фосфоглюкомутазы, фосфофруктокиназы. Болезнь Гирке. Гликогеноз - группа наслед. Заболеваний, харктеризующихся отложением в тканях либо большим кол-вом гликогена, либо необычным его видом. Сущ-ет 7 типов болезней гликогеноза: 1тип - болезнь Гирке- в печени, почках много гликогена, отсутствует повышение уровня глюкозы в крови. В печени, почках, кишечнике либо отсутствует либо низкое содержание глюкозо-6-фосфотазы. 2тип - болезнь Помпа- отсуствует глюкозидаза- она предотвращает накопление в лизосомах. 3тип - болезнь Форбса или Кори. 4тип - болезнь Андерсена- наступает летальный исход от печеночной недостаточности в первый год жизни. 5тип - синдром МакАрдля- невыносливость к физич. нагрузкам, в крови почти нету лактата. 6тип - недостаточ. фосфорилазы в печени. 7тип - недостаточ. фосфофруктокиназы в печени.
63. Сахарный диабет: причины, типы, сущность нарушений углеводного, липидного, белкового обменов. Принципы диагностики и лечения, осложнения. Сах.диабет – это хрон. заболевание, приводящее к нарушениям углеводного, белкового и липидного обмена в результате недостатка гормона инсулина или неправильного его действия. Сахарный диабет бывает первичным или вторичным: 1.Первичный (идиопатический) подразделен на: а)потенциальный; б)латентный проявляется. при лечении картизоном, острых инфекциях, беременности; характерен диабетический тип кривой сахарной нагрузки; в) асимптоматический (химический); г) клинический - делится на: «первого типа» инсулинзависимый -проявляется в характеризуется резким снижением секреции инсулина вследствие атрофии бета-клеток и «второго типа» инсулин-независимый. характеризуется некоторым снижением секреции инсулина. 2.Вторичный. Причины: а)повреждение поджелуд.ж-зы при хроническом панкреатите, опухолях и др. заболеваниях; б)присутствие антагонистов инсулина: избыточная секреция гормона роста или глюкокартикоидов; Другие причины диабета: 1.частичное нарушение нормальной структуры инсулина; 2.повыш. уровень секреции инсулиназы; 3. частичное нарушение структуры инсулиновых рецепторов.Различают 2 группы симптомов: основные и второстепенные. К основным симптомам относятся: Полиурия — усиленное выделение мочи, вызванное повышением осмотического давления мочи за счёт растворённой в ней глюкозы. Полидипсия (постоянная неутолимая жажда) — обусловлена значительными потерями воды с мочой и повышением осмотического давления крови. Полифагия — постоянный неутолимый голод, вызван нарушением обмена веществ при диабете, неспособностью клеток поглощать и перерабатывать глюкозу в отсутствие инсулина. Похудание - обусловлено повышенным катаболизмом белков и жиров из-за выключения глюкозы из энергетического обмена клеток. К вторичным симптомам относятся: зуд кожи и слизистых оболочек, сухость во рту, общая мышечная слабость, головная боль, нарушение зрения. Осложнения: Диабетический кетоацидоз — тяжёлое состояние, развивающееся вследствие накопления в крови продуктов промежуточного метаболизма жиров. Возникает при— инфекциях, травмах, операциях, при недостаточном питании. Гипогликемия — снижение уровня глюкозы в крови ниже нормального значения (обычно ниже 3,3 ммоль/л), происходит из-за передозировки сахароснижающих препаратов, сопутствующих заболеваний или недостаточного питании, приёма крепкого алкоголя. Гиперосмолярная кома, Встречается у пожилых больных с диабетом 2-го типа в анамнезе или без него и всегда связана с сильным обезвоживанием. Лактацидотическая кома - обусловлена накоплением в крови молочной кислоты и чаще возникает на фоне сердечно-сосудистой, печеночной и почечной недостаточности, как следствие этого, накопления в тканях молочной кислоты. Основной причиной развития лактацидотической комы является резкое смещение кислотно-основного равновесия в кислую сторону; обезвоживания, как правило, при этом виде комы не наблюдается.
64. Галактоземия. Причины. Сущность проявления заболевания. Галактоземия - редкое генетическое нарушение обмена веществ, при котором изменяется нормальный процесс метаболизма углеводов (сахаров) галактозы. Причины возникновения. При нормальном метаболизме, лактоза, содержащаяся в продуктах питания, под действием фермента лактазы, расщепляется, образуя глюкозу и галактозу. У людей, больных галактоземией, ферменты, необходимые для преобразования галактозы или отсутствуют, или же их уровень очень низок, что приводит к накоплению токсического галактозо-1-фосфата в различных тканях, эти процессы приводят к гепатомегалии (увеличение печени), циррозу печени, почечной недостаточности, катаракте, повреждениям головного мозга и яичников. Без лечения, смертность детей грудного возраста с диагнозом галактоземия составляет около 75%.
65. Переваривание и всасыв липидов в ЖКТ. транспорт в кровотоке. Пища ч-ка содержит простые липиды, слож.липиды, а также эфиры холестерола (ацетат, олеат, пальментат и стеорат). Переваривание липидов осущ-ся в тонком кишечнике при помощи желчи и панкреатического сока, у детей вырабатывается фермент слюны-язычная липаза, которая активна в желудке. В желудке липиды подвергаются кислому гидролизу,в результате чего образуется некоторое кол-во свобод. жирных кислот.При поступлении в тонкий кишечник происходит нейтрализация пищи при помощи бикарбонатов панкреатического сока. Далее в результате действия липаз происходит дальнейшее расщепление липидов. Желчные кислоты эмульгируют липиды, резко увеличивает площадь поверхности контакта водной и жировой фазы. Это необходимо, потому что липазы водорастворимы. При их действии образуется: свободные жиров.к-ты, лизолицетины, свободный холестерол.
66. Классы липопротеинов, их состав и ф-ции в транспорте липидов. Перенос триацилглицерола и холестерола в клетки. Липопротеины — сложные соединения, молекулы которых построены из липидов и белков, связанных между собой посредством гидрофобных и электростатических взаимодействий. Они входят в состав всех живых организмов, выполняют ф-цию транспорта и запасания липидов. Основные липопротеины плазмы: хиломикроны (образуются в клетках кишечника, богаты триглицеридами), липопротеины очень низкой (являются транспортной формой эндогенных триглицеридов), промежуточной, низкой (наиболее атерогенные липопротеины крови) и высокой плотности (самые мелкие, образуются в печени и кишечнике в виде незрелых дисковидных частиц). Холестерол поступает в клетки из крови в основном в составе лпнп, поступает в ткани путем эндоцитоза ЛП, некоторое количество холестерола поступает в клетки путём диффузии из ЛПНП и других ЛП при их контакте с мембранами клеток.
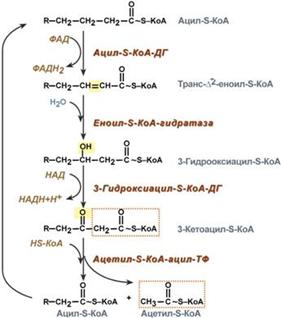
67. β-окисление жирных кислот. Окисление ненасыщенных жирных кислот с четным и нечетным числом углеродных атомов. Попадая в клетки триглицнриды подвергаются действию эстераз, которые отщипляют жирные кислоты. Образуется глицерол фосфорилируется превращается в дигидроксиацетонфосфат и включается в гликолиз. Жирные кислоты из цитоплазмы перемещаются в митохондрии, где происходит их расщепление. В зависимости от необходимости клетки отщепляют одно, двух-, трех углеродные фрагменты. Но в большинстве случаев клетке требуется клетке 2-х углеродный фрагмент. Перемещение жирных кислот осуществляется в три этапа: рис 1.

Далее в матриксе осуществляется расщепление жирных кислоты на 2-х углеродные фрагменты: рис 2. Окисление ненасыщенных жирной кислоты включает 2 этапа: 1. Выравнивание конфигурации за счет переноса группы СО из цис- в трансположение. 2. Гидратация присоединение OH-группы по 2-й связи и далее обычно. Окисление жирной кислоты с нечетным числом атомов углерода, обычное расщепление 2-х углеродных фрагментов дает в конце пропионил СоА, он карбоксилируется и изомеризуется, происходит внутримолекулярная перестройка –образуется сукцинил СоА.
68. НАРУШЕНИЯ ОБМЕНА ЛИПИДОВ Патологические состояния, связанные с отклонениями от нормального режима протекания метаболических процессов с участием липидов, вызываются, в первую очередь, нарушениями их переваривания и всасывания в организме. Первым симптомом нарушения обмена липидов является стеанорея, т.е. появление липидов в кале. Различают 3 основных типа стеанореи: 1) панкреатогеннаястеанорея, которая обусловлена низкой скоростью синтеза панкреатической липазы, что, в свою очередь, приводит к снижению интенсивности гидролиза триацилглицеринов в кишечном соке; 2) гепатогенная стеанорея, которая связана с нарушением поступления желчи в двенадцатиперстную кишку, в результате чего не происходит эмульгирования жиров, а, следовательно, и их гидролиза липазой; 3) энтерогенная стеанорея, которая обусловлена снижением метаболической активности слизистой оболочки тонкого отдела кишечника, в котором происходит ресинтез липидов.Увелич-е концентрации липопротеинов в крови приводит к возникновению гиперлипопротеинемии, а понижение - к гипопротеинемии. Гиперлипопротеинемии обусловлены замедлением расщепления липопротеиновых комплексов вследствие недостаточности липопротеинлипазы или в результате гиперинсулинизма, индуцирующего в печени усиленный синтез триацилглицеринов из углеводов. Гиперлипопротеинемия наряду с гиперхолестеролемией (увеличением содержания в крови холестерина) является основной причиной атеросклероза. Гиполипопротеинемии связаны как с нарушением переваривания и всасывания жиров в тонком отделе кишечника в результате дефицита липазы, а также с нарушениями продукции и поступления желчи. Кроме того, увелич-е содерж-я тиреоидных гормонов (гипертиреоз) приводит к усилению катаболизма сывороточных липидов.При атеросклерозе (отложение холестерина в стенках артерий) в стенках сосудов (преимущественно крупных артерий) откладываются жиры и прорастает соединительная ткань. Такие очаги разрешения сосудов называются атеросклеротическими бляшками. Атеросклеротические бляшки являются причиной сужения просвета сосудов и деформации участка кровеносного сосуда, что в свою очередь приводит к нарушениям циркуляции крови во внутренних органах. Сфинголипидозы - врожденные нарушения метаболизма липидов, главным образомсфинголипидов, обусловленные отсутствием лизосомных ферментов, катализирующих процессы распада сфинголипидов.Основу молекуулярной структуры сфинголипидов составляет церамид - продукт соединения через аминогруппу аминоспиртасфингозина и жирной кислоты. Разнообразие сфинголипидов связано с присоединением к церамиду более простых молекулярных групп, главным образом гексоз.Сфинголипиды являются важнейшими структурными компонентами клеточных мембран, в частности миелиновых оболочек нервных волокон, поэтому нарушение постоянно протекающего в организме их обновления, их распада в лизосомах клеток создает патологическую картину поражения большинства жизненно важных органов, включая серое и белое вещества головного мозга. Дефекты деградации сфинголипидов связаны с недостаточностью соответствующих ферментов специфических для каждого типа сфинголипидов.
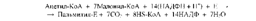 69. БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ. Биосинтез жирных к-т и липидов играет важную роль в жизнед-тиорг-мов. Именно в виде жирных к-т и триацилглицериновотклад-ся основные кол-ва энерг-х ресурсов орг-мов животных, в то время как энергоресурсы, откладываемые в форме углеводов, незначительны.В клетках орг-зма жирные к-ты синтез-тся из ацетил-КоА, образующегося из избыточной глюкозы пищи, которая не была использована организмом на энергетические нужды. В качестве восстановителя в биосинтезе жирных к-т принимает участие НАДФН, синтезируемый, в основном, в пентофосфатном пути распада углеводов. Нужно отметить, что хотя все реакции β-окисления жирных кислот обратимы, этот путь не используется организмом с целью их синтеза. Биосинтез жирных кислот осуществляется в цитоплазме клеток и катализируется целым полиферментным надмолекулярным ансамблем - пальмитилсинтетазой, состоящей из семи ферментов.Суммарная реакция биосинтеза жирных кислот в цитоплазме имеет следующий вид (Е - пальмитилсинтетаза):
69. БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ. Биосинтез жирных к-т и липидов играет важную роль в жизнед-тиорг-мов. Именно в виде жирных к-т и триацилглицериновотклад-ся основные кол-ва энерг-х ресурсов орг-мов животных, в то время как энергоресурсы, откладываемые в форме углеводов, незначительны.В клетках орг-зма жирные к-ты синтез-тся из ацетил-КоА, образующегося из избыточной глюкозы пищи, которая не была использована организмом на энергетические нужды. В качестве восстановителя в биосинтезе жирных к-т принимает участие НАДФН, синтезируемый, в основном, в пентофосфатном пути распада углеводов. Нужно отметить, что хотя все реакции β-окисления жирных кислот обратимы, этот путь не используется организмом с целью их синтеза. Биосинтез жирных кислот осуществляется в цитоплазме клеток и катализируется целым полиферментным надмолекулярным ансамблем - пальмитилсинтетазой, состоящей из семи ферментов.Суммарная реакция биосинтеза жирных кислот в цитоплазме имеет следующий вид (Е - пальмитилсинтетаза):
Из данного уравнения можно видеть, что для синтеза жирной кислоты требуется всего одна молекула ацетил-КоА, служащая «затравкой». Непосредственным источником синтеза является малонил-КоА, который образуется из ацетил-КоА по реакции:
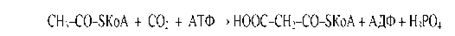
Эта реакция катализируется биотинзависимым ферментом - ацетил-КоА- карбоксилазой. Функция биотина сводится к переносу диоксида углерода на субстрат.Пальмитилсинтетаза представляет собой многофункциональный ансамбль белков: в центре полиферментного ансамбля находится ацилпереносящий белок (АПБ), содержащий свободную SH-группу; шесть остальных ферментов располагаются по периметру, причем один из них также содержит SH- группу. Поэтому пальмитилсинтетазу можно обозначить как:
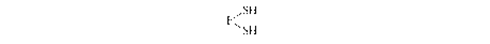
Процесс синтеза жирной кислоты описывается рядом последовательных реакций:1. Перенос ацетила с ацетил-КоА на синтетазу:

2. Перенос малонила с малонил-КоА на синтетазу:

3. Конденсация ацетила с малонилом и декарбоксилирование образовавшегося продукта:

4. Первое восстановление промежуточного продукта с участием НАДФН:
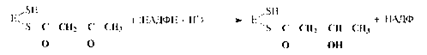
5. Дегидратация промежуточного продукта:
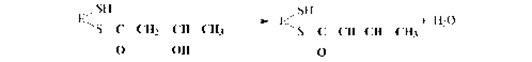
6. Второе восстановление промежуточного продукта с участием НАДФН:
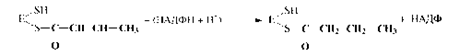
Затем синтезированный бутирил перемещается на ту SH-группу, с которой был связан затравочный ацетил, а на освободившуюся SH-группу поступает новый малонильный остаток из малонил-КоА. Далее цикл повторяется снова; после семи оборотов цикла синтезируется пальмитил-Е, который при участии пальмитилдеацилазыгидролизуется до пальмитиновой кислоты и фермента (Е). Пальмитиновая кислота - это основной продукт биосинтеза, однако в небольших количествах могут образовываться и другие жирные кислоты.Жирные кислоты с разветвленной углеродной цепью синтезируются из продуктов метаболизма аминокислот с разветвленной цепью (валин, изолейцин и лейцин) через ацильные производные КоА путем удлинения цепи и при участии АПБ. Особенности биосинтеза полиненасыщенных жирных кислот представляют интерес в связи с их витаминоподобными функциями. Некоторые полиеновые кислоты могут синтезироваться из олеиновой кислоты с помощью ряда последовательных реакций. Однако, синтез полиненасыщенных кислот, содержащих двойные связи, расположенные между конечным метилом и седьмым атомом углерода, невозможен, поэтому они и являются незаменимыми в пищевом рационе.Т.о., биосинтез и поступление с пищей - два основных источника жирных кислот для организма человека и животных. Биосинтез триацилглицеринов. Образующиеся в результате биосинтеза жирные кислоты в организмах животных и человека в свободном виде встречаются лишь в незначительных количествах, а присутствуют главным образом в виде триацилглицеринов. Синтез триацилглицеринов происходит в печени и жировой ткани из КоА-производных жирных кислот через фосфатидную кислоту по реакции:
Не нашли, что искали? Воспользуйтесь поиском: