ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
ПОПУЛЯЦИОННЫЕ ВОЛНЫ
Популяционными волнами или волнами жизни (С.С. Четвериков) называют периодические или апериодические колебания численности организмов в природных популяциях. Это явление распространяется на все виды животных и растений, а также на микроорганизмы. Причины колебаний часто имеют экологическую природу. Так, размеры популяций «жертвы» (зайца) растут при снижении давления на них со стороны популяций «хищника» (рыси, лисицы, волка). Отмечаемое в этом случае увеличение кормовых ресурсов способствует росту численности хищников, что, в свою очередь, интенсифицирует истребление жертвы (рис. 11.1).
Вспышки численности организмов некоторых видов, наблюдавшиеся в ряде регионов мира, были обусловлены деятельностью человека.В XIX—XX вв. это относится к популяциям кроликов в Австралии, домовых воробьев в Северной Америке, канадской элодеи в Евразии. В настоящее время существенно возросли размеры популяций домовой мухи, находящей прекрасную кормовую базу в виде разлагающихся пищевых отбросов вблизи поселений человека. Напротив, численность 12 популяций домовых воробьев в городах падает вследствие прекращения широкого использования лошадей. Масштабы колебаний численности организмов разных видов варьируют. Для одной из зауральских популяций майских жуков отмечены изменения количества особей в 106 раз.

Рис. 11.1 Колебания численности особей в популяциях жертвы (заяц-беляк, сплошная линия) и хищников (А - рыси; B - лисицы; Д - волка)
Численность особей выражена в процентах к минимальному из зарегистрированных значений, принятому за 100%
Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. При росте численности организмов наблюдается слияние ранее разобщенных популяций и объединение их генофондов. Так как популяции по своему генетическому составу уникальны, в результате такого слияния возникают новые генофонды с измененными по сравнению с исходными частотами аллелей. В условиях возросшей численности интенсифицируются межпопуляционные миграции особей, что также способствует перераспределению аллелей. Рост количества организмов обычно сопровождается расширением занимаемой территории.
На гребне популяционной волны некоторые группы особей выселяются за пределы ареала вида и оказываются в необычных условиях существования. В таком случае они испытывают действие новых факторов естественного отбора. Повышение концентрации особей в связи с ростом их численности усиливает внутривидовую борьбу за существование.
При спаде численности наблюдается распад крупных популяций. Возникающие малочисленные популяции характеризуются измененными генофондами. В условиях массовой гибели организмов редкие мутантные аллели могут быть генофондом потеряны. При сохранении редкого аллеля его концентрация в генофонде малочисленной популяции автоматически возрастает. На спаде волны жизни часть популяций, как правило, небольших по размерам, остается за пределами обычного ареала вида. Чаще они, испытывая действие необычных условий жизни, вымирают. Реже, при благоприятном генетическом составе, такие популяции переживают период спада численности. Будучи изолированными от основной массы вида, существуя в необычной среде, они нередко являются родоначальниками новых видов.
Популяционные волны — это эффективный фактор преодоления генетической инертности природных популяций. Вместе с тем их действие на генофонды не является направленным. В силу этого они, так же как и мутационный процесс, подготавливают эволюционный материал к действию других элементарных эволюционных факторов.
ИЗОЛЯЦИЯ
Ограничение свободы скрещиваний (панмиксии) организмов называют изоляцией. Снижая уровень панмиксии, изоляция приводит к увеличению доли близкородственных скрещиваний. Сопутствующая этому гомозиготизация усиливает особенности генофондов популяций, которые создаются вследствие мутаций, комбинативной изменчивости, популяционных волн. Препятствуя снижению межпопуляционных генотипических различий, изоляция является необходимым условием сохранения, закрепления и распространения в популяциях генотипов повышенной жизнеспособности.
В зависимости от природы факторов ограничения панмиксии различают географическую, биологическую и генетическую изоляцию. Географическая изоляция заключается в пространственном разобщении популяций благодаря особенностям ландшафта в пределах ареала вида — наличию водных преград для «сухопутных» организмов, участков суши для видов-гидробионтов, чередованию возвышенных участков и равнин. Ей способствует малоподвижный или неподвижный (у растений) образ жизни. Так, на Гавайских о-вах популяции наземных улиток занимают долины, разделенные невысокими гребнями. Сухость почвы и редколесье затрудняют преодоление этих гребней моллюсками. Выраженная, хотя и неполная, изоляция в течение многих поколений привела к ощутимым различиям фенотипов улиток из разных долин. В горах о-ва Оаху, например, один из видов улиток Achatinella mustelina представлен более чем сотней рас, выделяемых по морфологическим признакам.
Пространственная изоляция может происходить и в отсутствие видимых географических барьеров. Причины ее в таком случае кроются в ограниченных «радиусах индивидуальной активности». Так, у «береговой» рыбы бельдюги Zoarces viviparus от устья у концу фьорда уменьшается число позвонков и лучей некоторых плавников. Сохранение изменчивости объясняется оседлым образом жизни бельдюги. Такая изменчивость наблюдается и у подвижных видов животных, например перелетных птиц с гнездовым консерватизмом. Молодь ласточек, например, возвращается с зимовки на место своего рождения и гнездится в радиусе до 2 км от материнского гнезда. Скрещивания у ласточек ограничиваются группой близко селящихся особей. В отличие от разделения барьерами эту разновидность географической изоляции обозначают как разделение расстоянием.
Биологическая изоляция возникает вследствие внутривидовых различий организмов и имеет несколько форм. К экологической изоляции приводят особенности окраски покровов или состава пищи, размножение в разные сезоны, у паразитов — использование в качестве хозяина организмов разных видов. Так, в Молдове есть две несмешивающиеся популяции мышей —желтогорлая лесная мышь и степная. Фактором разделения их является состав пищи. Разобщение популяций способствовало выявлению и усилению особенностей фенотипа степных мышей. Они мельче и имеют иную форму черепа. В описанном примере экологическая изоляция дополняется территориальной. Сезонные расы, выделяемые по месту и времени икрометания, описаны у лососевых, осетровых, карповых рыб.
Длительная экологическая изоляция способствует дивергенции популяций вплоть до образования новых видов. Так, предполагают, что человеческая и свиная аскариды, морфологически очень близкие, произошли от общего предка. Их расхождению, согласно одной из гипотез, способствовал запрет на употребление человеком в пищу свиного мяса, который по религиозным соображениям распространялся длительное время на значительные массы людей.
Этологическая (поведенческая) изоляция существует благодаря особенностям ритуала ухаживания, окраски, запахов, «пения» самок и самцов из разных популяций. Так, подвиды щеглов — седоголовый Carduelis carduelis carduelis и черноголовый С. с. brevirostis — имеют выраженные отметины на голове. Серые вороны Corvus corone cornix из крымской и североукраинской популяций, внешне неразличимые, отличаются карканьем.
При физической (механической) изоляции препятствием к скрещиванию являются различия в структуре органов размножения или просто разница в размерах тела. У растений такая форма изоляции возникает при приспособлении цветка к определенному виду опылителей.
Описанные формы изоляции, особенно в начальный период их действия, снижают, но не исключают полностью межпопуляционные скрещивания.
Генетическая (репродуктивная) изоляция создает более жесткие, иногда непреодолимые барьеры скрещиваниям. Она заключается в несовместимости гамет, гибели зигот непосредственно после оплодотворения, стерильности или малой жизнеспособности гибридов.
Иногда разделение популяции сразу начинается с генетической изоляции. К этому приводят полиплоидия или массивные хромосомные перестройки, резко изменяющие хромосомные наборы гамет мутантов по сравнению с исходными формами. Полиплоидия распространена среди растений (рис. 11.2). Разные виды плодовой мухи нередко различаются хромосомными перестройками. Гибриды от скрещивания близкородственных форм со сниженной жизнеспособностью известны для серой и черной ворон. Указанный фактор изолирует популяции этих птиц в Евразии (рис. 11.3). Чаще генетическая изоляция развивается вторично вследствие углубления морфологических различий организмов из популяций, длительно разобщенных другими формами изоляции — географической, биологической. В первом случае генетическая изоляция предшествует дивергенции признаков и начинает процесс видообразования, во втором — она его завершает.


Рис.11.2. Растения и наборы хромосом в соматических клетках Solanum nigrum:
а—г: 36, 72, 108 и 144 хромосомы
Изоляция в процессе видообразования взаимодействует с другими элементарными эволюционными факторами. Она усиливает генотипические различия, создаваемые мутационным процессом и генетической комбинаторикой. Возникающие благодаря изоляции внутривидовые группировки отличаются по генетическому составу и испытывают неодинаковое давление отбора.
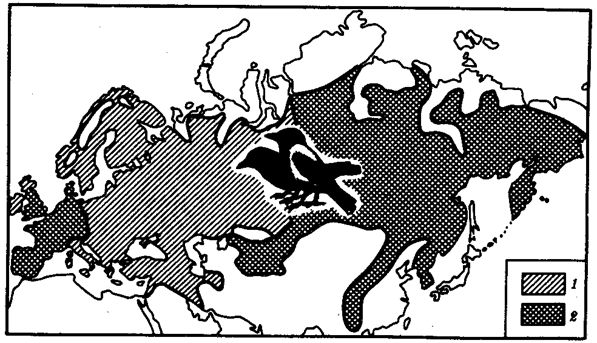
Рис. 11.3. Сниженная жизнеспособность гибридов как фактор разделения популяций серой и черной ворон:
1 — ареал серой вороны, 2 — ареал черной вороны
ЕСТЕСТВЕННЫЙ ОТБОР
В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Благодаря индивидуальной изменчивости в условиях конкретной среды обитания приспособленность разных генотипов (фенотипов) различна. В эволюционном контексте приспособленность определяют как произведение жизнеспособности в данной среде, обусловливающей большую или меньшую вероятность достижения репродуктивного возраста, на репродуктивную способность особи. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции.
Необходимой предпосылкой отбора является борьба за существование — конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная (избирательная) смертность. В конечном итоге отбор обеспечивает дифференциальное (избирательное) воспроизведение (размножение) генотипов. Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов.
Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия, отдельные особи — объектами действия, а конкретные признаки — точками приложения отбора.
Эффективность отбора по качественному и количественному изменению генофонда популяции зависит от величины давления и направления его действия. Величину давления отбора выражают коэффициентом отбора S, который характеризует интенсивность устранения из репродуктивного процесса или сохранения в нем соответственно менее или более приспособленных форм по сравнению с формой, принятой за стандарт приспособленности. Так, если некий локус представлен аллелями A1 и А2, то популяция по генотипам делится на три группы: A1A1; A1A2; A2A2. Обозначим приспособленность этих генотипов W0, W1, W2. Выберем в качестве стандарта первый генотип, относительная приспособленность которого максимальна и равна 1. Тогда приспособленности других генотипов составят доли этого стандарта:
AlA1 A1A2 A2A2
W0 W1 W2
или W0/W0 = 1, W1/W0=l—S1, W2/W0=l—S2.
Величины S1 и S2 означают пропорциональное снижение в очередном поколении воспроизводства генотипов A1A2 и А2А2 сравнительно с генотипом AlA1.
Отбор особенно эффективен в отношении доминантных аллелей при условии их полного фенотипического проявления и менее эффективен в отношении рецессивных аллелей, а также в условиях неполной пенетрантности. На результат отбора влияет исходная концентрация аллеля в генофонде. При низких и высоких концентрациях отбор происходит медленно. Изменение доли доминантного аллеля в сравнении с рецессивным при коэффициенте отбора 0,01 приведено ниже.
| Возрастание, % | Число поколений |
| 0,1-1 | |
| 1-50 | |
| 50-98 | |
| 98-99,9 |
В теории, упрощая ситуацию, допускают, что отбор через фенотипы действует на генотипы благодаря различиям в приспособительной ценности отдельных аллелей. В реальной жизни приспособительная ценность генотипов зависит от влияния на фенотип и взаимодействия всей совокупности генов. Оценка величины давления отбора по изменению концентрации отдельных аллелей технически часто невыполнима. Поэтому расчет проводят по изменению концентрации организмов определенного фенотипа.
Пусть в популяции присутствуют организмы двух фенотипических классов А и В в отношении СA/СB = U1. Вследствие различий по приспособленности происходит естественный отбор (селекция), который изменяет соотношение особей с фенотипами А и В. В следующем поколении оно станет СA/СB = U2 = U1 (1 + S), где S — коэффициент отбора. Отсюда S = U2/U1 — 1. При селективном преимуществе фенотипа А U2 > U1, a S > 0. При селективном преимуществе фенотипа В U2 < U1 и S < 0. Если приспособленность фенотипов А и В сопоставима и U2 = U1, a S=0. В рассмотренном примере при S > 0 отбор сохраняет в популяции в ряду поколений фенотипы А и устраняет фенотипы В, при S < 0 имеет место обратная тенденция. Отбор, сохраняющий определенные фенотипы, по своему направлению является положительным, тогда как отбор, устраняющий фенотипы из популяции,— отрицательным.
В зависимости от результата различают стабилизирующую, движущую и дизруптивную формы естественного отбора (рис. 11.4). Стабилизирующий отбор сохраняет в популяции средний вариант фенотипа или признака. Он устраняет из репродуктивного процесса фенотипы, уклоняющиеся от сложившейся адаптивной «нормы», приводит к преимущественному размножению типичных организмов. Так, сотрудник одного из университетов США подобрал после снегопада и сильного ветра 136 оглушенных воробьев Passer domesticus. Из них 72 выживших воробья имели крылья средней длины, тогда как 64 погибшие птицы были либо длиннокрылыми, либо короткокрылыми. Стабилизирующая форма соответствует консервативной роли естественного отбора. При относительном постоянстве условий среды благодаря этой форме сохраняются результаты предшествующих этапов эволюции.
Движущий (направленный) отбор обусловливает последовательное изменение фенотипа в определенном направлении, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. При смене условий обитания благодаря этой форме отбора в популяции закрепляется фенотип, более соответствующий среде. После того как новое значение признака придет в оптимальное соответствие условиям среды, движущая форма отбора сменяется стабилизирующей. Примером такого отбора является замещение в популяции гавани Плимут (Англия) крабов Carcinus maenas с широким головогрудным щитком животными с узким щитком в связи с увеличением количества ила.

Рис. 11.4. Формы естественного отбора:
I —стабилизирующая, II— движущая, III— дизруптивная;
F1—F3— последовательные поколения особей

Рис. 11.5. Изменение массы тела мышей из двух популяций в последовательных поколениях вследствие отбора по этому признаку
1 — тяжелые животные, 2 — легкие животные
Направленный отбор составляет основу искусственного отбора. Так, в одном эксперименте на протяжении ряда поколений из популяции шестинедельных мышей отбирали для скрещивания наиболее тяжелых и наиболее легких животных. Избирательное воспроизведение по признаку массы тела привело к образованию двух самостоятельных популяций, соответственно с возрастающей и убывающей массой тела (рис. 11.5). По окончании опыта, занявшего 11 поколений, ни одна из этих популяций не вернулась к первоначальной массе.
Дизруптивный (разрывающий) отбор сохраняет несколько разных фенотипов с равной приспособленностью. Он действует против особей со средним или промежуточным значением признаков. Так, в зависимости от преобладающего цвета почвы улитки Cepaea nemoralis имеют раковины коричневой, желтой, розовой окраски. Дизруптивная форма отбора «разрывает» популяцию по определенному признаку на несколько групп. Она поддерживает в популяции состояние генетического полиморфизма.
В зависимости от формы отбор сокращает масштабы изменчивости, создает новую или сохраняет прежнюю картину разнообразия. Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения соотношений аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направленны. Отбор приводит генофонды в соответствие с критерием приспособленности. Он осуществляет обратную связь между изменениями генофонда и условиями обитания, накладывает на эти изменения печать биологической целесообразности (полезности). Естественный отбор действует совместно с другими эволюционными факторами. Поддерживая генотипическое разнообразие особей в ряду поколений, мутационный процесс, а также популяционные волны, комбинативная изменчивость создают для него необходимый материал.
Естественный отбор нельзя рассматривать как «сито», сортирующее генотипы по приспособленности. В эволюции ему принадлежит творческая роль. Исключая из размножения генотипы с малой приспособительной ценностью, сохраняя благоприятные генные комбинации разного масштаба, он преобразует картину генотипической изменчивости, складывающуюся первоначально под действием случайных факторов, в биологически целесообразном направлении. Результатом творческой роли отбора является процесс органической эволюции, идущей в целом по линии прогрессивного усложнения морфофизиологической организации (арогенез), а в отдельных ветвях — пути специализации (аллогенез).
Не нашли, что искали? Воспользуйтесь поиском: