ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Пасивний транспорт іонів
У відсутності градієнта концентрації перенос заряджених частинок (іонів) може відбуватися при наявності електричного поля, тобто градієнта електричного потенціалу 
Густина електричного струму іонів за законом Ома
 (4.19)
(4.19)
де  - коефіцієнт електропровідності середовища. Як відомо, густина електричного струму
- коефіцієнт електропровідності середовища. Як відомо, густина електричного струму
 (4.20)
(4.20)
де  - кількість носіїв заряду (іонів) в одиниці об'єму середовища;
- кількість носіїв заряду (іонів) в одиниці об'єму середовища;  - швидкість, з якою рухаються носії (іони) під впливом електричного поля;
- швидкість, з якою рухаються носії (іони) під впливом електричного поля;  - заряд носія (іона).
- заряд носія (іона).
Оскільки  ' - рухливість іона, а
' - рухливість іона, а 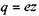 , де
, де  - валентність іона, то вираз для густини струму набуває такого вигляду:
- валентність іона, то вираз для густини струму набуває такого вигляду:
 (4.21)
(4.21)
Згідно з визначенням густини струму
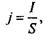 (4.22)
(4.22)
де
 (4.23)
(4.23)
Заряд 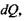 що проходить через площину
що проходить через площину  за час
за час 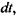 задовольняє такому співвідношенню:
задовольняє такому співвідношенню:
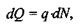 (4.24)
(4.24)
де 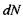 - кількість іонів, що перетинають площину
- кількість іонів, що перетинають площину  за час
за час  Об'єднавши вирази (4.22), (4.23) і (4.24), одержимо
Об'єднавши вирази (4.22), (4.23) і (4.24), одержимо
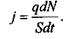 (4.25)
(4.25)
Згідно з визначенням потоку речовини, маємо
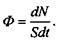 (4.26)
(4.26)
Порівнюючи (4.25) і (4.26), одержимо такий вираз для потоку іонів, зумовленого градієнтом електричного потенціалу:
 (4.27)
(4.27)
Рухливість іона пов'язана з коефіцієнтом дифузії співвідношенням Ейнштейна

де  - число Фарадея;
- число Фарадея;  - абсолютна температура;
- абсолютна температура;  - газова стала. Тоді рівняння (4.27) набуває вигляду
- газова стала. Тоді рівняння (4.27) набуває вигляду

При наявності як градієнта концентрації іонів, так і градієнта потенціалу електричного поля, потік іонів
 (4.28)
(4.28)
Рівняння (4.28) відоме як електродифузійне рівняння Нернста-Планка, що описує пасивний транспорт заряджених частинок (іонів).
Виконавши деякі елементарні перетворення, рівняння Нернста-Планка можна записати таким чином:
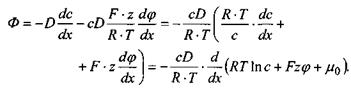 (4.29)
(4.29)
Тут враховано ту обставину, що 
Крім того, під знак похідної (градієнта) внесено постійну величину - хімічний потенціал розчинника 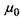 , який є однаковим по обидва боки мембрани і тому не залежить від координати, по якій виконується диференціювання. Величина, яка стоїть у дужках, тобто
, який є однаковим по обидва боки мембрани і тому не залежить від координати, по якій виконується диференціювання. Величина, яка стоїть у дужках, тобто 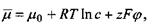
називається електрохімічним потенціалом  . Його фізичний зміст полягає в тому, що він визначає вільну енергію Гібса з розрахунку на один моль у присутності розчиненої речовини і електричного поля. Другий доданок
. Його фізичний зміст полягає в тому, що він визначає вільну енергію Гібса з розрахунку на один моль у присутності розчиненої речовини і електричного поля. Другий доданок  визначає внесок розчиненої речовини (осмотичний внесок) в електрохімічний потенціал, у той час як останній доданок
визначає внесок розчиненої речовини (осмотичний внесок) в електрохімічний потенціал, у той час як останній доданок  характеризує внесок електричного поля. Враховуючи означення електрохімічного потенціалу, рівняння (4.29) можна подати у такому вигляді:
характеризує внесок електричного поля. Враховуючи означення електрохімічного потенціалу, рівняння (4.29) можна подати у такому вигляді:
 (4.30)
(4.30)
Рівняння (4.30) відоме як рівняння Теорелла. Згідно з ним рушійною силою пасивного транспорту іонів виступає градієнт електрохімічного потенціалу, у той час як рушійною силою пасивного транспорту незаряджених молекул є градієнт концентрації.
Процес простої (або звичайної) дифузії відбувається повільно і слабо контролюється клітиною. За таким механізмом здійснюється транспорт кисню, вуглекислого газу та шкідливих для клітини речовин (наприклад, ядів). При звичайній дифузії молекула дифундуючої речовини рухається крізь мембрану без утворення комплексів з іншими молекулами.
Для більш швидкого переносу речовин, необхідних для життєдіяльності клітин, в ході еволюції виробились інші типи пасивної дифузії: перенос через канали (пори) і перенос за допомогою переносників. Ці типи дифузії відомі під назвою полегшеної дифузії (рис.4.26).

Рис. 4.26. Види пасивного і активного транспорту через мембрану: 1 - проста дифузія через мембрану; 2 - дифузія через канали; 3 - дифузія за допомогою переносників; 4 - активний транспорт.
Розглянемо більш детально пасивний транспорт речовин крізь канали. Канали - це білкові утворення, що мають діаметр  Згідно з деякими даними білкові канали покривають
Згідно з деякими даними білкові канали покривають 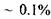 поверхні мембрани. Слід підкреслити, що канали - не статичні, а динамічні утворення. Один з механізмів їх утворення - процес латеральної дифузії. Коефіцієнт проникності мембрани при пасивному транспорті через канали
поверхні мембрани. Слід підкреслити, що канали - не статичні, а динамічні утворення. Один з механізмів їх утворення - процес латеральної дифузії. Коефіцієнт проникності мембрани при пасивному транспорті через канали

де  - кількість каналів на одиницю площі мембрани;
- кількість каналів на одиницю площі мембрани; 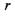 - радіус каналу;
- радіус каналу;  - довжина каналу (товщина мембрани);
- довжина каналу (товщина мембрани);  - коефіцієнт дифузії речовини у воді. Згідно з гіпотезою Мулінза найкраще проходять крізь канали ті іони, радіус яких в оточенні одного шару молекул води близький до радіуса каналу.
- коефіцієнт дифузії речовини у воді. Згідно з гіпотезою Мулінза найкраще проходять крізь канали ті іони, радіус яких в оточенні одного шару молекул води близький до радіуса каналу.
Швидкість пасивного транспорту крізь мембрани збільшується на кілька порядків у присутності переносників -молекул, що володіють дуже високим рівнем селективності. Наприклад, переносник, який полегшує транспорт глюкози крізь мембрану, ніяк не впливає на транспорт амінокислот. Найбільш детально полегшений транспорт крізь біологічні мембрани було вивчено на прикладі переносу іонів іонофорними антибіотиками типу валіноміцину. Було встановлено, що валіноміцин вибірково збільшує проникність мембран для іонів 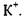 Шість карбонільних груп валіноміцину формують порожнину, в яку потрапляє іон
Шість карбонільних груп валіноміцину формують порожнину, в яку потрапляє іон  У вільній молекулі валіноміцину атоми складноефірних карбонільних груп спрямовані до зовнішньої поверхні молекули. При утворенні комплексу з іонами
У вільній молекулі валіноміцину атоми складноефірних карбонільних груп спрямовані до зовнішньої поверхні молекули. При утворенні комплексу з іонами  у молекулі виникають додаткові іон-дипольні взаємодії з атомами кисню карбонільних груп. Молекула набуває форми браслета діаметром близько
у молекулі виникають додаткові іон-дипольні взаємодії з атомами кисню карбонільних груп. Молекула набуває форми браслета діаметром близько  у центрі якого розташовується іон
у центрі якого розташовується іон  На рис. 6.27 показано конформаційну зміну молекули валіноміцину при створенні комплексу з іоном
На рис. 6.27 показано конформаційну зміну молекули валіноміцину при створенні комплексу з іоном  Іони
Іони  надто великі для порожнини у молекулі валіноміцину, внаслідок чого різниця у проникностях для іонів
надто великі для порожнини у молекулі валіноміцину, внаслідок чого різниця у проникностях для іонів  становить:
становить:  Таким чином, основою селективності при переносі іонів за допомогою переносників, так само, як і при переносі через канали, є принцип структурної відповідності.
Таким чином, основою селективності при переносі іонів за допомогою переносників, так само, як і при переносі через канали, є принцип структурної відповідності.
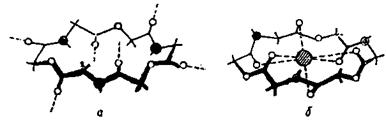
Рис. 4.27. Просторова структура переносника іонів К+: а - валіноміцину; б - його комплексу з іоном.
Транспорт за допомогою переносників може відбуватись внаслідок дифузії переносника разом з речовиною (рухомий переносник) (рис. 4.28а), а також і шляхом естафетної передачі від однієї молекули переносника до іншої (рис. 4.28б).
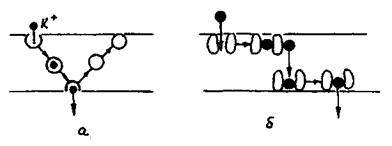
Рис. 4.28. Транспорт іонів за участю переносників: а - рухомий переносник; б - естафетний перенос.
Для пасивного транспорту за допомогою переносників характерний ефект концентраційного насичення швидкості переносу: зі збільшенням концентрації швидкість переносу сповільнюється, тому що всі переносники виявляються пов'язаними з певною частиною іонів.
Активний транспорт
Поряд з пасивним транспортом у життєдіяльності клітини важливу роль відіграє активний транспорт - примусовий перенос молекул та іонів з області малих концентрацій до області високих концентрацій. Завдяки активному транспорту підтримуються концентраційні градієнти, які необхідні для нормального функціонування клітини. Транспортні системи, які створюють необхідні концентраційні градієнти, називають насосами, або АТФазами. Відомі чотири основні системи активного транспорту:
1.  2.
2.  3.
3. 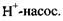 4. Перенос протонів під час роботи дихального ланцюга мітохондрій. Активний транспорт, як і пасивний, забезпечується спеціальними структурами: каналами, переносниками, ферментами. При активному транспорті (на відміну від пасивного) вектор переміщення іонів співпадає за напрямком з вектором концентраційного градієнта, тобто з напрямком збільшення концентрації. Активний транспорт відбувається за рахунок енергії, що виділяється при гідролізі АТФ (комплексу
4. Перенос протонів під час роботи дихального ланцюга мітохондрій. Активний транспорт, як і пасивний, забезпечується спеціальними структурами: каналами, переносниками, ферментами. При активному транспорті (на відміну від пасивного) вектор переміщення іонів співпадає за напрямком з вектором концентраційного градієнта, тобто з напрямком збільшення концентрації. Активний транспорт відбувається за рахунок енергії, що виділяється при гідролізі АТФ (комплексу  ) з утворенням молекул АДФ і неорганічного фосфату
) з утворенням молекул АДФ і неорганічного фосфату  Гідроліз АТФ здійснюється ферментом АТФазою. Джерелом молекул АТФ є процеси окислювального фосфорилювання, що відбуваються в мітохондріях.
Гідроліз АТФ здійснюється ферментом АТФазою. Джерелом молекул АТФ є процеси окислювального фосфорилювання, що відбуваються в мітохондріях.
Розглянемо основні етапи роботи найбільш дослідженого 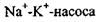 (рис. 4.29). Як відомо, в нормально функціонуючій клітині концентрація іонів
(рис. 4.29). Як відомо, в нормально функціонуючій клітині концентрація іонів  перевищує їх концентрацію в міжклітинному середовищі, а для іонів
перевищує їх концентрацію в міжклітинному середовищі, а для іонів 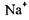 це співвідношення обернене. Наприклад, у нервовому волокні кальмара:
це співвідношення обернене. Наприклад, у нервовому волокні кальмара:

 локалізована у зовнішніх плазматичних мембранах клітин. Через те, що
локалізована у зовнішніх плазматичних мембранах клітин. Через те, що  є гідрофобним білком, вона створює комплекси з ліпідами бішару. Саме тому активність інтегрального білка
є гідрофобним білком, вона створює комплекси з ліпідами бішару. Саме тому активність інтегрального білка  АТФази залежить від фізико-хімічного стану мембрани. Встановлено, що для функціонування
АТФази залежить від фізико-хімічного стану мембрани. Встановлено, що для функціонування 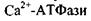 важливо, щоб ліпідне оточення було рідким, на відміну від
важливо, щоб ліпідне оточення було рідким, на відміну від 
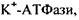 для якої важлива впорядкованість бішару.
для якої важлива впорядкованість бішару.
Рис. 4.29. Основні етапи роботи  насоса.
насоса.
На першому етапі відбувається приєднання до АТФ-ази трьох іонів 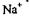 і фосфорилювання ферменту.
і фосфорилювання ферменту.
На другому етапі - перенос центрів зв'язування  зовні (транслокація І).
зовні (транслокація І).
На третьому етапі - відокремлення трьох іонів  і заміна їх двома іонами
і заміна їх двома іонами 
На четвертому етапі - відщеплення залишки фосфорної кислоти.
На п'ятому етапі - перенос центрів зв'язування іонів  всередину клітини (транслокація II).
всередину клітини (транслокація II).
На шостому етапі - відщеплення двох іонів  і приєднання трьох іонів
і приєднання трьох іонів  та фосфорилювання ферменту (тобто повторюється етап І).
та фосфорилювання ферменту (тобто повторюється етап І).

Перенос двох іонів 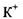 всередину клітини і трьох іонів
всередину клітини і трьох іонів 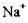 зовні спричиняє перенос одного позитивного заряду із цитоплазми в навколишнє середовище. Внаслідок цього всередині клітини виникає негативний електричний потенціал. Тому
зовні спричиняє перенос одного позитивного заряду із цитоплазми в навколишнє середовище. Внаслідок цього всередині клітини виникає негативний електричний потенціал. Тому 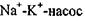 називають електрогенним. Таким чином, при активному транспорті потік іонів
називають електрогенним. Таким чином, при активному транспорті потік іонів  співпадає з напрямками градієнта концентрації
співпадає з напрямками градієнта концентрації  і градієнта електричного потенціалу
і градієнта електричного потенціалу  а потік іонів
а потік іонів 
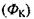 - напрямлений по градієнту концентрації
- напрямлений по градієнту концентрації 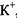 , але проти
, але проти  (рис. 4.30).
(рис. 4.30).
Не нашли, что искали? Воспользуйтесь поиском: