ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Оптимальные температуры для выращивания растений
| Растение | Температура, °С | |
| Дневная | Ночная | |
| Фиалка африканская Петуния Цинния Левкой Маргаритка Астра Томаты Эшшольция |
Температурный оптимум большинства живых организмов находится в пределах 20—25 С, и лишь у обитателей жарких, сухих районов температурный оптимум жизнедеятельности находится несколько выше 25—28°С. Например, некоторые прямокрылые (кузнечики) проявляют полуденную активность в условиях пустыней Палестины при температуре 40°С и выше.
Для организмов умеренных и холодных зон России оптимальные температуры от 10 до 20°С. Так, у ветреницы дубравной процесс фотосинтеза наиболее интенсивно протекает при 10°С.
В зависимости от ширины интервала температуры, в которой данный вид может существовать, организмы делятся на эвритермные и стенотермные. Эвритермные организмы выдерживают широкие колебания температуры, стенотермные живут лишь в узких пределах.
К эвритермным относится большинство организмов районов с континентальным климатом. Многие из них имеют покоящие стадии, переносящие особенно широкий диапазон температуры (покоящиеся яйца, цисты, куколки насекомых, находящиеся в состоянии анабиоза, взрослые животные, споры бактерий, семена растений).
Беспозвоночные, рыбы, амфибии и рептилии лишены способности поддерживать температуру тела в узких границах. Их называют пойкилотермными (от греч. poikilos — разный). Данных животных часто называют также эктотермными, так как они больше зависят от тепла, поступающего извне, чем от того тепла, которое образуется в обменных процессах. Характерна низкая интенсивность обмена и отсутствие механизма сохранения тепла. Раньше этих животных обычно называли холодокровными, но этот термин неточен и может вводить в заблуждение.
Птицы и млекопитающие способны поддерживать достаточно постоянную температуру тела независимо от окружающей температуры. Этих животных называют гомойотермными (от греч. homoios — подобный) или, по старой терминологии, что менее правильно, теплокровными. Гомойотермные животные относительно мало зависят от внешних источников тепла. Благодаря высокой интенсивности обмена у них вырабатывается достаточное количество тепла, которое может сохраняться. Поскольку эти животные существуют за счет внутренних источников тепла, их называют в настоящее время чаще эндотермными.
Растения и животные в ходе длительного эволюционного развития, приспосабливаясь к периодическим изменениям температурных условий, выработали в себе различную потребность к теплу в разные периоды жизни. Например, прорастание семян растений происходит при более низких температурах, чем последующий их рост. Семена пшеницы, овса, ячменя прорастают при 1—2°С, всходы же появляются при 4—5^0. В период цветения растениям, как правило, необходимо больше тепла, чем в период созревания семян, плодов. Томаты лучше растут и развиваются, когда температура днем 25—26 °С, ночью 17—18°С. Температурный оптимум живых организмов зависит и от других экологических факторов. Установлено, что при полном освещении и избытке углекислого газа в воздухе оптимальная температура фотосинтеза 30 °С, а при слабом освещении и недостатке углекислого газа она снижается до 10°С (рис. 4.8).
При характеристике температуры необходимо различать температуру воздуха и температуру почвы, разность между ними. Для растений это особенно важно, так как они способны поглощать питательные вещества из почвы при условии, если температура почвы будет на несколько градусов ниже температуры воздуха. Например, гречиха достигает наилучшего развития, когда температура близ корней равнг. 10°С, а у надземных частей 22°С. При температуре почвы и воздуха 22°С состояние растений резко ухудшается, и они не дают цветков. При дальнейшем повышении температуры почвы до 34°С, когда надземные органы остаются при 22°С, у растений наблюдается отмирание верхушек почек, стеблей, а впоследствии погибает все растение.

Рис. 4.8. Соотношение между фотосинтезом и дыханием
в зависимости от температур
При оптимальных температурах у всех организмов физиологические процессы протекают наиболее интенсивно, что способствует увеличению темпов их роста. Здесь к биологическим процессам вполне приемлемо правило Вант-Гоффа (Т.А. Акимова, В.В. Хаскин, 1998).
Так, если скорость Vт реакции измерена при двух температурах Т1 и Т2, причем Т1 < Т2, то температурный коэффициент Вант-Гоффа:

Зависимость скорости реакции от температуры может быть выражена уравнением Аррениуса:
 (4.2)
(4.2)
где АV — фактор частоты событий, называемый также константой Аррениуса;
Е* — энергия активации данной реакции (Дж/моль), необходимая для преодоления потенциального барьера реакции;
R — газовая постоянная [8,3144 Дж/(моль. К)];
Т — абсолютная температура, К.
В диапазоне температур 15 — 40 °С (288—313 К) значения Q,g большинства биохимических процессов лежат между 1,5 и 2,5, а значения Е* — между 30 и 65 кДж/моль.
Исходя из этого правила скорость химических реакций возрастает в 2—3 раза при повышении температуры на каждые 10°С. При температурах выше или ниже оптимальных скорость биохимических реакций в организме снижается или вообще нарушается. И как итог — замедление темпов роста и даже гибель организма.
В пределах от верхних оптимальных до верхних максимальных и от нижних минимальных до нижних оптимальных температур лежат диапазоны верхнего и нижнего пессимумов. Развитие растений при температурном пессимуме осуществляется замедленными темпами и затягивается на длительное время.
Активность животных также ограничивается пессимумами. У насекомых повышение температуры вызывает вначале медленные, некоординированные движения, в физиологической области (оптимум) приводит к полностью управляемой активности, а при дальнейшем повышении — к чрезмерно быстрым, некоординированным, суматошным движениям. Так, муха цеце при температуре ниже 8°С неподвижна, при 10°С начинает бегать, выше 14°С при дополнительном раздражении взлетает, а выше 21°С летает сонливо.
Температурный оптимум разных видов и стадий развития у насекомых также неодинаков. Например, оптимальная температура развития яиц озимой совки (Agrotis segetum) 25°C, гусениц 22 °С, а куколок 19°С.
Крайне минимальные и максимальные температуры нижнего и верхнего пессимумов называются соответственно нижним и верхним порогом развития, или нижним и верхним биологическим нулем, за пределами которого развитие организма не происходит.
Температуры, лежащие выше нижнего порога развития и не выходящие за пределы верхнего, получили название эффективных температур. Для растений и эктотермных животных количество тепла, необходимого для развития, определяется суммой эффективных температур или суммой тепла. Зная нижний порог развития, легко определить эффективную температуру — по разности наблюдаемой и пороговой температур. Так, если нижний порог развития организма равен 10°С, а реальная в данный момент температура воздуха 25°C, то эффективная температура будет 15°С (25°—10°). Сумма эффективных температур определяется по формуле:
C = (t –t1)×n, (4.3)
где С — сумма эффективных температур;
t — температура окружающей среды (реальная, наблюдаемая);
t1 — температура порога развития;
n — продолжительность (длительность) развития в днях, часах.
Сумма эффективных температур для каждого вида растений и эктотермных животных, как правило, величина постоянная, при том, что если другие условия среды находятся в оптимуме, отсутствуют осложняющие факторы. При отклонении этих условий или при сравнении особей из разных частей ареала результаты могут быть искажены. Например, в Северо-Западном регионе России цветение мать-и-мачехи начинается при сумме эффективных температур равной 77, кислицы — 453, земляники — 500, желтой акации — 700°С. Ограничивающим фактором географического расположения видов нередко является сумма эффективных температур, которую нужно набрать для завершения жизненного цикла. Так, северная граница древесной растительности в целом совпадает с июльскими изотермами 10, 12°С. Севернее уже не хватает тепла для развития деревьев, и зона лесов сменяется безлесными тундрами.
Развитие эндотермных животных в меньшей степени зависит от температуры окружающей среды. И тем не менее и им свойствен определенный температурный оптимум и пессимум тех или иных физиологических процессов.
У крупного рогатого скота повышение температуры в помещениях при их содержании до 15 °С или понижение до 7 °С приводит к снижению плодовитости.
Живые организмы в процессе эволюции выработали различные формы адаптации к температуре, среди них морфологические, биохимические, физиологические, поведенческие и т. д. Растения не имеют собственной температуры тела и по отношению к тепловому фактору обладают определенной спецификой. Одно из важнейших приспособлений к температуре у растений — форма их роста. Там, где тепла мало — в Арктике, в высокогорье, много подушковидных растений, растений с прикорневыми розетками листьев, стелющихся форм. Так, у стланцевых форм карликовой березы, ели, можжевельника и кедровника верхние ветви, поднимающиеся высоко над землей, большей частью полумертвые или мертвые, а стелющиеся — живые, так как зимуют под снегом и не подвергаются отрицательному воздействию низких температур. Все это позволяет растениям улавливать максимум тепла солнечных лучей, а также использовать тепло нагретой поверхности почвы (рис.4.9).

(из Д. П. Шенникова,1950)
Температурный фактор на развитие приземистых форм растений может действовать как непосредственно, так и косвенно, вызывая нарушения водоснабжения и минерального питания.
Наиболее значительна роль прямого влияния температур в процесса геофилизации растений (рис. 4.10).
Геофилизация — это погружение базальной (нижней) части растения в почву — сначала гипокотиля, затем эпикотиля, первого междоузлия и т. д. Это характерно преимущественно покрытосеменным растениям. Геофилизация в ходе их исторического развития играла значительную роль в трансформации жизненных форм от деревьев до трав.

Рис. 4.10. Геофилизация (втягивание в почву) подсемядольного
колена клевера лугового (Trifolium pratense), по П. Лисицину:
а — поверхность почвы; б — глубина втягивания
Сильные холода и чрезвычайная жара нередко ограничены во времени, и растения избегают их воздействия, сбрасывая чувствительные части, или редуцируют свое вегетативное тело до подземных многолетних органов. При наступлении благоприятных условий они вновь образуют надземные органы. Здесь важно знать и устойчивость к температуре различных органов с учетом их функций. Особенно чувствительны к низким температурам (холоду) репродуктивные органы — зачатки цветков в зимующих почках и завязи в цветках (рис. 4.11).
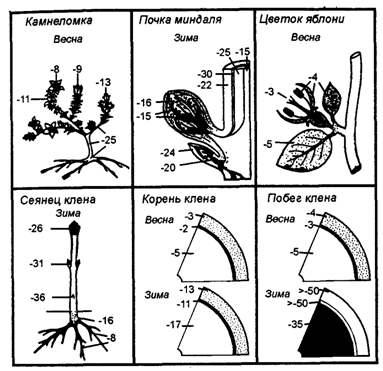
Рис. 4.11. Холодостойкость отдельных органов
и тканей разных растений (по В. Лархеру, 1978)
При распространении растений необходимо учитывать устойчивость цветков в почках, самих цветков, семян и незащищенных молодых растений или наиболее чувствительных стадий развития, которые большей частью ограничивают сохранение и расселение вида, так называемое правило Тинеманна.
Распространена у растений жарких мест способность впадать в состояние вынужденного покоя.
У животных морфологические адаптации к температуре прослеживаются четко. Под действием теплового фактора у животных формируются такие морфологические признаки, как отражательная поверхность тела, пуховой, перьевой и шерстный покровы у птиц и млекопитающих, жировые отложения. Большинство насекомых в Арктике и высоко в горах имеет темную окраску. Это способствует усиленному поглощению солнечного тепла. Темный пигмент яиц многих водных животных выполняет ту же функцию. Эндотермные животные, обитающие в холодных областях (полярные медведи, киты и др.), имеют, как правило, крупные размеры, тогда как обитатели жарких стран (например, многие насекомоядные млекопитающие) обычно меньше по размерам. Это явление носит название правила Бергмана. Согласно этому правилу, при продвижении на север средние размеры тела в популяциях эндотермных животных увеличиваются (табл. 4.7).
Таблица 4.7
Изменение размера тела животных с широтой (по Бергману)
| Вид | Район | Длина тела, см | Масса, кг |
| Волк Лиса | Таймыр Монголия Среднерусская равнина Туркмения | До 137 До 120 До 90 До 57 | До 49 До 40 До 10 До 3,2 |
При увеличении размеров уменьшается относительная поверхность тела, а следовательно, и теплоотдача.
Размеры выступающих частей тела также варьируют в соответствии с температурой среды. У видов, живущих в более холодном климате, различные выступающие части тела (хвост, уши, конечности и др.) меньше, чем у родственных видов из более теплых мест. Это явление известно как правило Аллена. Правило Аллена наглядно проявляется при сравнении длины ушей у трех видов лисиц, обитающих в разных географических областях рис. 4.12).
 Арктический вид
Арктический вид
Температура тела 37°С
Средняя температура среды 0°С
Европейский вид
Температура тела 37°С
Средняя температура среды 12°С
Африканский вид
Температура тела 37°С
Средняя температура среды 25°С
Рис. 4.12. Различия в длине ушей у трех видов лисиц,
обитающих в разных географических областях
(по Н. Грину и др., 1993)
Третье правило (носит название правила Глогера) гласит, что окраска животных в холодном и сухом климате сравнительно светлее, чем в теплом и влажном. Эти правила (часто их называют законами), управляющие адаптациями млекопитающих, равным образом относятся и к человеку.
Биохимическая адаптация живых организмов к температуре проявляется прежде всего в изменении физико-химического состояния веществ, содержащихся в клетках и тканях. Так, при адаптации к низким температурам в клетках растения благодаря увеличению запаса пластических веществ повышается концентрация растворов, увеличивается осмотическое давление клеточного сока, уменьшается содержание свободной воды, не связанной в коллоиды. И это очень важно, так как «связанная» вода трудно испаряется и замерзает, слабо отжимается под давлением, обладает большой плотностью и в значительной степени утрачивает свойство растворителя. Она становится кристаллической по структуре и в то же время сохраняет жидкое состояние. Между частицами цитоплазмы и водой устанавливается единство структуры, обеспечивающее ей таким образом вхождение в структуру макромолекул белков и нуклеиновых кислот. В таком состоянии ее трудно заморозить, перевести в твердое состояние. Важным приспособлением к низким температурам является и отложение запасных питательных веществ в виде высокоэнергетических соединений — жира, масла, гликогена и др. Так, И.М. Васильев (1970) описал значение отложения запасных веществ в растении в форме масла. Он утверждает, что масло прежде всего вытесняет воду из вакуоли и этим предохраняет растительный организм от замерзания. Масло, откладываясь в цитоплазме, делает ее более стойкой к морозу и к другим неблагоприятным воздействиям зимнего периода. Такую же роль играют откладываемые в протоплазму и вакуоли крахмал и белки. Большое значение имеют и те биохимические изменения в запасных питательных веществах, которые протекают в период подготовки к зимнему состоянию. Так, значительная часть накопленного в летний период крахмала вновь превращается в сахар. При этом появляются сахара, которых обычно мало содержится в клетках летом. Например, зимой в клетках тканей коры у хвойных помимо сахарозы, глюкозы и фруктозы появляются стахиоза и рафиноза. В летний период они содержатся в других частях растения.
К тканевым механизмам приспособления к действию низких температур относится своеобразное распределение резервных энергетических веществ в теле организмов. При адаптации к холоду, по данным исследований, у организмов происходит «перемещение» веществ в органах. У тех или иных видов растений нередко к зиме масла и сахара откладываются в тканях надземных органов, а в подземных органах — крахмал. При этом в районах с очень низкими температурами у растений отмечается значительное накопление масла во внутренних слоях древесины, что повышает их устойчивость к сильным морозам. У животных, и в первую очередь обитателей полярных областей, с понижением температуры возрастает содержание гликогена в печени, повышается содержание аскорбиновой кислоты в тканях почек. У млекопитающих большое скопление питательных веществ наблюдается в бурой жировой ткани в непосредственной близости от жизненно важных органов — сердца и спинного мозга — и имеет приспособительный характер. В митохондриях клеток этой ткани при клеточном дыхании не синтезируется АТФ, а вся энергия рассеивается в виде тепла.
Многие животные к зиме накапливают жир. Подкожный жировой слой обеспечивает им теплоизоляцию. У ряда животных в выступающих или поверхностных частях тела (лапы некоторых птиц, ласты китов) есть замечательное приспособление под названием «чудесная сеть». Это сплетение сосудов, в котором вены тесно прижаты к артериям. Кровь, текущая по артериям, отдает тепло венам, оно возвращается к телу, а артериальная кровь поступает в конечности охлажденной. Конечности, по существу, пойкилотермны, зато температуру остального тела можно поддерживать с меньшими затратами энергии. На основе физиологических процессов многие организмы способны в определенных пределах менять температуру своего тела. Эта способность называется терморегуляцией. Как правило, терморегуляция сводится к тому, что температура тела поддерживается на более постоянном уровне по сравнению с температурой окружающей среды. Особенно совершенны механизмы терморегуляции у эндотермных животных. Как уже было отмечено ранее, эндотерм-ные животные способны вырабатывать достаточное количество тепла и регулировать теплоотдачу, поэтому равенство прихода и расхода тепла сохраняется (рис. 4.13).
Система терморегуляции млекопитающих и птиц включает рецепторы, эффекторы и чрезвычайно чувствительный регуляторный центр в гипоталамусе. Этот центр следит за температурой крови, отражающей температуру тех органов, через которые она протекает.

Рис.4.13. Схема теплообмена между телом лошади (температура 38°С) и окружающей средой в жаркий солнечный день при температуре воздуха 30°С. Прерывистыми линиями показана передача тепла путем излучения
Поддерживать температуру тела на постоянном уровне животным помогает испарение жидкости с поверхности тела при высоких температурах окружающей среды. У человека для этого служит потоотделение, у собак и многих птиц — учащенное дыхание. Некоторые сумчатые в жару обмазывают шкуру обильной слюной.
Пути теплообмена между пойкилотермным организмом и окружающей средой показаны на рис. 4.14.

Рис. 4.14. Схематическое изображение путей теплообмена между
пойкилотермным организмом и окружающей средой
(по Е. А. Криксунову и др., 1995)
Среди пойкилотермных животных некоторые также способны к терморегуляции при определенных условиях. Шмели, бражники, крупные вараны, отдельные виды рыб, например тунцы, могут повышать температуру тела в периоды высокой мышечной активности.
У животных есть разнообразные поведенческие адаптации к температуре. Они проявляются в перемещениях животных в места с более благоприятными температурами (перелеты, миграции), в изменениях сроков активности, сдвигая ее на более светлое время суток и т. д. В пустыне, где днем поверхность почвы может нагреваться до 60—70 °С, на раскаленном песке животных почти не увидишь. Насекомые, рептилии и млекопитающие проводят жаркое время, зарывшись в песок или спрятавшись в норы. В глубине почвы температура не так резко колеблется и сравнительно невысокая. Холодным утром кузнечики подставляют бока солнечному свету, а дневные бабочки расправляют крылья. В полуденную жару они, сложив крылья, располагаются параллельно лучам.
При понижении температуры воздуха многие животные переходят на питание более калорийной пищей. Белки в теплое время года поедают более ста видов кормов, зимой же питаются главным образом семенами хвойных, богатых жирами. Кормом оленям летом в основном служат травы, зимой — лишайники, содержащие в большом количестве белковые, жировые и сахаристые вещества.
Важное место в преодолении отрицательного воздействия низких температур, особенно в зимний период, занимает выбор животными места для жилища, утепление убежищ, гнезд пухом, сухими листьями, углубление нор, закрывание входов в них, принятие особой позы (например, скручивание кольцом, укутывание хвостом), собирание в группы, так называемое «скучивание» и т.д. Некоторые животные согреваются путем пробежек и прыжков.
При всем многообразии приспособлений живых организмов к воздействию неблагоприятных температурных условий среды выделяют три основных пути: активный, пассивный и избегание неблагоприятных температурных воздействий.
Активный путь — усиление сопротивляемости, развитие ре-гуляторных способностей, дающих возможность осуществления жизненных функций организма, несмотря на отклонения температур от оптимума. Этот путь ярко выражен у эндотермных животных, развит у эктотермных, в зачаточной форме проявляется у некоторых высших растений. Пассивный путь — это подчинение жизненных функций организма ходу внешних температур. Недостаток тепла вызывает угнетение жизнедеятельности, что способствует экономному использованию энергетических запасов. И как итог — повышение устойчивости клеток и тканей организма. Данный путь приспособления к воздействию неблагоприятных температур характерен для всех растений и эктотермных животных. Элементы пассивного приспособления, или адаптации, присущи и эндотермным животным, обитающим в условиях крайне низких температур. Выражается это в снижении уровня обмена, замедлении скорости роста и развития, позволяющее экономнее расходовать ресурсы в сравнении с быстро развивающимися видами. У млекопитающих и птиц преимущества пассивного приспособления в неблагоприятные периоды года используют гетеротермные виды, которые обладают способностью впадать в спячку или оцепенение.
Избегание неблагоприятных температурных воздействий — общий способ для всех организмов. Выработка жизненных циклов, когда наиболее уязвимые стадии развития проходят в самые по температурным условиям благоприятные периоды года. Для растений это главным образом изменения в ростовых процессах, для животных — разнообразные формы поведения.
В связи с тем что растения и животные исторически приспособлены к определенным тепловым режимам, совершенно закономерно, что температурный фактор имеет непосредственное отношение к их распределению на Земле и обусловливает в той или иной мере заселенность природных зон живыми организмами. Одной из главных закономерностей в распределении современных организмов является их биополярность. Она заключается в том, что у организмов в высоких широтах умеренных зон наблюдается определенное сходство в систематическом составе и ряде биологических явлений. Это характерно как для наземной, так и для морской фауны и флоры. Биополярность отмечается и в поширотном качественном составе живых организмов. Например, для тропической зоны характерно более высокое видовое разнообразие по сравнению с высокими широтами.
Влажность
Вода. В жизни организмов вода выступает как важнейший экологический фактор. Без воды нет жизни. Живых организмов, не содержащих воду, на Земле не найдено. Она является основной частью протоплазмы клеток, тканей, растительных и животных соков. Все биохимические процессы ассимиляции и диссимиляции, газообмен в организме осуществляются при наличии воды. Вода с растворенными в ней веществами обусловливает осмотическое давление клеточных и тканевых жидкостей, включая и межклеточный обмен. В период активной жизнедеятельности растений и животных содержание воды в их организмах, как правило, довольно высокое (табл. 4.8).
Таблица 4.8
Содержание воды в растительных и животных организмах,
% к массе тела (по Б. С. Кубанцеву, 1973)
| Растения | Содержание воды | Животные | Содержание воды |
| Водоросли Корни моркови Листья трав Листья деревьев Клубни картофеля Стволы деревьев | 96-98 87-91 83-86 79-82 74-80 40-55 | Губки Моллюски Насекомые Ланцетник Земноводные Млекопитающие | 80-92 46-92 До 93 68-83 |
В недеятельном состоянии организма количество воды может значительно снижаться, однако и в период покоя она не исчезает полностью. Например, в сухих мхах и лишайниках содержится воды 5—7% к общей массе, а в воздушно-сухих зерновках злаков — не менее 12—14%. Наземные организмы из-за постоянной потери воды нуждаются в регулярном ее пополнении. Поэтому у них в процессе эволюции выработались приспособления, которые регулируют водный обмен и обеспечивают экономное расходование влаги. Приспособления носят анатомо-морфологический, физиологический и поведенческий характер. Потребность разных видов растений в воде по периодам развития не одинакова. Меняется она и в зависимости от климата и почвы. Так, злаковые культуры в периоды прорастания семян и созревания нуждаются меньше во влаге, чем во время наиболее интенсивного роста. Кроме влажных тропиков, практически повсеместно растения испытывают временный недостаток воды, засуху. При высоких температурах в летний период часто проявляется атмосферная засуха, почвенная — при уменьшении доступной растению почвенной влаги. Недостаток или дефицит влаги снижает прирост растений, может стать причиной их низкорослости, бесплодия из-за недоразвития генеративных органов. Первостепенное значение во всех проявлениях жизнедеятельности имеет водный обмен между организмом и внешней средой. Влажность среды нередко является фактором, лимитирующим распространение и численность организмов на Земле. Например, степные и особенно лесные растения требуют повышенного содержания паров в воздухе, растения же пустынь приспособились к низкой влажности.
Важную роль у животных играют проницаемость покровов и механизмы, регулирующие водный обмен. Здесь уместно дать характеристику основным показателям влажности. Влажность — это параметр, характеризующий содержание водяного пара (газообразной воды) в воздухе. Различают абсолютную и относительную влажность. Абсолютная влажность — количество газообразной воды, содержащейся в воздухе, и выраженное через массу воды на единицу массы воздуха (например, в граммах на 1 кг или на 1 м3 воздуха). Относительная влажность — это отношение количества имеющегося в воздухе пара к насыщенному количеству пара при данных условиях температуры и давления. Это соотношение устанавливается по формуле:
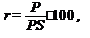 (4.4)
(4.4)
где r — относительная влажность;
Р и PS — абсолютная и насыщающая (максимальная) влажность при данной температуре.
Относительную влажность обычно измеряют, сравнивая температуру на двух термометрах — с влажным и сухим шариком. Этот прибор называется психрометром. Так, если оба термометра показывают одинаковую температуру, то относительная влажность равна 100%. Если же «влажный» термометр показывает меньшую температуру, чем «сухой» (обычно так и бывает), то и относительная влажность будет меньше 100%. Точную величину получают из специальных справочных таблиц. Для измерения относительной влажности удобен и гигрограф. В приборе используется свойство человеческого волоса сокращаться или удлиняться в зависимости от относительной влажности, что позволяет вести непрерывную запись показаний.
В экологических исследованиях относительная влажность измеряется довольно часто. Большое значение для организмов имеет и дефицит насыщения воздуха водяными парами или разность между максимальной и абсолютной влажностью при определенной температуре. Дефицит насыщения воздуха можно обозначить буквой и определить по формуле:
d = PS - Р. (4.5)
Этот показатель наиболее четко характеризует испаряющую силу воздуха и играет для экологических исследований особую роль. В связи с тем, что испаряющая сила воздуха с повышением температуры увеличивается, при разных температурах дефицит насыщения неодинаков при одной и той же влажности. При его возрастании воздух становится суше и в нем интенсивнее происходят испарение и транспирация. При уменьшении дефицита насыщения относительная влажность воздуха увеличивается. Температура среды самым существенным образом влияет на характер действия влажности.
Важными в жизни организмов являются и особенности распределения влаги по сезонам в течение года. Выпадают ли осадки в зимнее или летнее время? Каково суточное ее колебание? Так, в северных районах нашей планеты обильные осадки, выпадающие в холодное время года, большей частью недоступны растениям, и в то же время даже малые осадки летом оказываются жизненно необходимыми. Важно учитывать и характер выпадающих осадков — моросящий дождь, ливень, снег, их продолжительность. Например, моросящий дождь летом хорошо увлажняет почву, более эффективен для растений, чем ливень, несущий колоссальные потоки воды. Во время ливня почва не успевает впитывать воду, она быстро стекает, унося с собой плодородную часть, плохо укоренившиеся растения, зачастую ведет к гибели мелких животных, особенно насекомых. Однако и затянувшиеся моросящие дожди могут оказывать неблагоприятное воздействие на жизнедеятельность животных, например насекомоядных птиц в период выкармливания птенцов.
Зимние осадки, выпадающие в виде снега в холодном и умеренном климате, создают снежный покров, который благоприятно влияет на температурный режим почвы и тем самым повышает выживаемость растений и животных. И наоборот, зимние осадки в виде дождя оказывают неблагоприятное воздействие на растения, их выживаемость, увеличивают смертность насекомых.
Степень насыщения воздуха и почвы водяными парами имеет большое значение. Нередко наблюдаются случаи гибели животных и растений во время засухи, которая вызвана чрезмерной сухостью воздуха или суховеями. В первую очередь это сказывается на жизни организмов, обитающих во влажных местах, как правило, из-за отсутствия у них механизмов, регулирующих потерю воды при транспирации и испарении, тогда как наружные покровы тела весьма непроницаемы.
Влажность воздуха обусловливает периодичность активной жизни организмов, сезонную динамику жизненных циклов, влияет на продолжительность развития, плодовитость и их смертность. Например, такие виды растений, как вероника весенняя, незабудка песчаная, бурачок пустынный и др., используя весеннюю влагу, успевают в очень короткие сроки (12—30 дней) прорасти, развить генеративные побеги, расцвести, сформировать плоды и семена. Данные однолетние растения называют эфемерами (от греч. ephemeros — мимолетный, однодневный). Эфемеры, в свою очередь, подразделяются на весенние и осенние. Вышеназванные растения относятся к весенним эфемерам. Четкую приспособленность к сезонному ритму влажности проявляют и отдельные виды многолетних растений, называемые эфемероидами или геоэфемероидами. При неблагоприятных условиях влажности они могут задерживать свое развитие до тех пор, пока она не станет оптимальной, или, как эфемеры, пройти весь его цикл в чрезвычайно сжатые ранневесенние сроки. Сюда можно отнести типичные растения южных степей — гиацинт степной, птицемлечники, тюльпаны и др.
Эфемерами могут быть и животные. Это такие, как насекомые, ракообразные (щитни, появляющиеся в большом количестве весной в лесных лужах), и даже рыбы, обитающие в небольших водоемах, лужах, например африканские нотобранхи и афиосемионы из отряда карпозубообразных.
По отношению к влажности различают эвригигробионтные и стеногигробионтные организмы. Эвригигробионтные организмы приспособились жить при различных колебаниях влажности. Для стеногигробионтных организмов влажность должна быть строго определенной: высокой, средней или низкой. Развитие животных не менее тесно связано с влажностью среды. Однако животные в отличие от растений имеют возможность активно отыскивать условия с оптимальной влажностью, обладают более совершенными механизмами регуляции водного обмена.
Влажность среды влияет на содержание воды в тканях животного и отсюда имеет непосредственное отношение к его поведению и выживаемости. Вместе с тем она может оказывать и косвенное воздействие через пищу и другие факторы. Например, во время засух при сильном выгорании растительности сокращается численность животных-фитофагов. Развитие животных по фазам требует строго определенных условий по влажности. При недостатке влаги в воздухе или пище у животных резко сокращается плодовитость, и в первую очередь у влаголюбивых форм. Недостаточное количество воды в корме снижает интенсивность роста у большинства животных, замедляет их развитие, сокращает продолжительность жизни, увеличивает смертность (рис. 4.15).

Рис. 4.15. Влияние влажности на основные жизненные
процессы у животных (по Н. П. Наумову, 1963):
А — гигрофилы; Б — ксерофилы;
1 — смертность; 2 — долговечность; 3 — плодовитость; 4 — скорость развития
Следовательно, водный режим, т. е. последовательные изменения в поступлении, состоянии и содержании воды во внешней среде (дождь, снег, туман, насыщение парами воздуха, уровень грунтовых вод, влажность почвы), оказывает существенное влияние на жизнедеятельность живых организмов.
По отношению к водному режиму наземные организмы подразделяются на три основные экологические группы: гигрофильные (влаголюбивые), ксеро-фильные (сухолюбивые) и мезофильные (предпочитающие умеренную влажность). Примером гигрофилов среди растений могут служить ка-лужница болотная, кислица обыкновенная, лютик ползучий, чистяк лютичный и др.; среди животных — мокрецы, ногохвостки, комары, стрекозы, жужелицы, уж и т. д. Все они не выносят значительного водного дефицита и плохо переносят даже кратковременную засуху.
Настоящими ксерофилами являются жуки-чернотелки, верблюды, вараны. Здесь широко представлены многообразные механизмы регуляции водного обмена и приспособления к удержанию воды в теле и клетках, что слабо выражено у гигрофилов.
Вместе с тем разделение организмов на три группы в какой-то мере относительно, так как у многих видов степень потребности во влаге непостоянна в различных условиях и неодинакова на разных стадиях развития организмов. Так, проростки и молодые растения многих древесных пород развиваются по типу мезофильных, а взрослые растения имеют явные черты ксерофилов.
По способу регулирования водного режима наземные растения подразделяются на две группы: пойкилогидридные и гомео-гидридные. Пойкилогидридные растения — это виды, не способные активно регулировать свой водный режим. У них нет каких-либо особенностей анатомического строения, которые способствовали бы защите от испарения. У большинства отсутствуют устьица. Транспирация равна простому испарению. Содержание воды в клетках находится в равновесии с давлением паров в воздухе или определяется его влажностью, зависит от его колебаний. К пойкилогидридным растениям относятся грибы, наземные водоросли, лишайники, некоторые мхи, из высших растений — тонколистные папоротники тропических лесов. Немногочисленную группу составляют цветковые растения, имеющие устьица, — представители семейства геснериевых, обитающие в расщелинах скал на Балканах и в Южной Африке. Сюда же относят среднеазиатскую пустынную осоку — Сагех physodes. Листья пойкилогидридных растений способны высыхать практически до воздушно-сухого состояния, а после смачивания вновь «оживают» и зеленеют.
Гомеогидридные растения способны в определенных пределах регулировать потерю воды путем закрывания устьиц и складывания листьев. В клеточных оболочках откладываются водонепроницаемые вещества (суберин, кутин), поверхность листьев покрыта кутикулой и т. д. Это дает возможность гомеогидридным растениям поддерживать на сравнительно постоянном уровне содержание воды в клетках и давление водяных паров в межклетниках. Транспирация по величине, дневной и сезонной динамике значительно отличается от свободного испарения смоченного физического тела (рис. 4.16).

Рис. 4.16. Схема суточного хода транспирации при
разной водообеспеченности растений (из Т. К. Горышиной, 1979):
1 — Транспирация без ограничения; 2 — Транспирация с полуденным снижением благодаря сужению устьиц; 3 — то же, при полном закрытии устьиц; 4 — полное исключение устьичной транспирации благодаря длительному закрыванию устьиц (остается лишь кутикулярная транспирация); 5 — снижение кутикулярной транспирации благодаря изменению проницаемости мембран. Стрелки, направленные вниз, — закрывание устьиц; стрелки, направленные вверх, — открывание устьиц. Пунктир — дневной ход испарения со свободной водной поверхности. Штриховка — область кутикулярной транспирации
Эта группа составляет большинство высших сосудистых растений, формирует растительный покров Земли. Иначе вместо зеленых лесов и лугов даже в умеренных широтах свежая зелень встречалась бы только после дождей.
Условия водного обмена у организмов определяются влажностью местообитания. В зависимости от этого у них появляются черты приспособления к жизни в условиях достаточного или малого водоснабжения. Наиболее четко это выражено у растений. Не обладая возможностью свободного передвижения, они лучше других проявляют приспособленность к жизни в местообитаниях с большим или малым количеством влаги.
В зависимости от местообитания среди наземных растений различают следующие экологические группы: гигрофиты, мезофиты и ксерофиты. Гигрофиты (от греч. «гигрос» — влажный и «фитон» — растение) — растения, обитающие во влажных местах, не переносящие водного дефицита и обладающие невысокой засухоустойчивостью. Растения этой группы имеют, как правило, крупные, тонкие, нежные листовые пластинки с небольшим числом устьиц, нередко расположенных с обеих сторон. Устьица большей частью широко открыты, в связи с этим транспирация мало отличается от физического испарения. Корни обычно толстые, слабо разветвленные. Корневые волоски представлены слабо или отсутствуют. Все органы покрыты тонким однослойным эпидермисом, кутикулы практически нет. Широко развита аэренхима (возду-хоносная ткань), обеспечивающая аэрацию тела растения. К гигрофитам в первую очередь относят тропические растения, которые живут при высокой температуре и влажности воздуха. Нередко гигрофиты обитают в тени под пологом леса (например, папоротники) или на открытых пространствах, но обязательно на почвах переувлажненных или покрытых водой. В умеренном и холодном климате типичными гигрофитами являются теневые травянистые растения лесов. На открытых местах и влажных почвах растут световые гигрофиты. Это такие, как калужница (Calta palustris), плакун-трава (Lythrum salicaria), росянка (Drosera), многие злаки и осоки сырых местообитаний, из культурных растений к световым гигрофитам относят рис, культивируемый на полях, залитых водой.
В целом же при довольно большом разнообразии местообитаний, особенностей водного режима и анатомоморфологичес-ких черт всех гигрофитов объединяет отсутствие приспособлений, ограничивающих расход воды и неспособность выносить даже незначительную ее потерю.
Например, у световых гигрофитов листья в дневное время могут терять за час количество воды, которое в 4—5 раз превышает массу листа. Хорошо известно, как быстро вянут в руках цветы, собранные по берегам водоемов. Показательны для гигрофитов и небольшие величины сублетального водного дефицита. Для кислицы и майника потеря 15—20% запаса воды уже необратима и ведет к гибели.
Мезофиты — это растения умеренно увлажненных местообитаний. Они имеют хорошо развитую корневую систему. На корнях имеются многочисленные корневые волоски. Листья разные по размеру, но, как правило, большие, мягкие, нетолстые, плоские, с умеренно развитыми покровной, проводящей, механической, столбчатой и губчатой тканями. Устьица располагаются на нижней стороне листовых пластинок. Хорошо выражена регуляция устьичной транспирации. К мезофитам относятся многие луговые травы (клевер луговой, тимофеевка, ежа сборная), большинство лесных растений (ландыш, зеленчук и др.), значительная часть лиственных деревьев (береза, осина, клен, липа), многие полевые (рожь, картофель, капуста) и плодово-ягодные (яблоня, смородина, вишня, малина) культуры и сорняки.
Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая во влажных условиях более гигроморфные, а в сухих — более ксероморфные черты.
Мезофиты связаны переходами с другими экологическими типами растений по отношению к воде, поэтому четкую границу между ними провести зачастую очень трудно. Например, среди луговых мезофитов выделяются виды с повышенным влаголюбием, предпочитающие постоянно сырые или временно заливаемые участки (лисохвост луговой, бекмания обыкновенная, канареечник тростниковый и др.). Их объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми лесными травами, предпочитающими лесные овраги, переувлажненные или наиболее сырые леса, такие, как селезеночник, недотрога, папоротник, некоторые лесные мхи и др.
В местообитаниях с периодическим или постоянным (но невысоким) недостатком влаги встречаются мезофиты с повышенной физиологической устойчивостью к засухе, с теми или иными ксе-роморфными признаками. Эта группа переходная между мезофитами и ксерофитами носит название ксеромезофиты. Сюда относят многие виды растений северных степей, сухих сосновых боров, песчаных местообитаний — клевер-белоголовку, подмаренник желтый и др., из культурных растений — люцерну, засухоустойчивые сорта пшеницы и некоторые другие. Ксерофиты (от греч. «ксерос» — сухой и «фитон» — растение) — это растения сухих местообитаний, способные переносить значительный недостаток влаги — почвенную и атмосферную засуху. Наиболее обильны и разнообразны ксерофиты в областях с жарким и сухим климатом. К ним принадлежат виды растений пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков и т. д.
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен ограниченным поступлением воды при ее недостатке в почве и увеличением расхода влаги на транспирацию при большой сухости воздуха и высокой температуре. Таким образом, для преодоления недостатка влаги могут быть разные пути: увеличение ее поглощения и сокращение расхода, а также способность переносить большие потери воды. При этом различают два основных способа преодоления засухи: возможность противостоять иссушению тканей, или активное регулирование водного баланса, и способность выносить сильное иссушение.
Важное значение для ксерофитов имеют разнообразные структурные приспособления к условиям недостатка влаги. Например, сильное развитие корневой системы помогает растениям увеличить поглощение почвенной влаги. Нередко у травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9—10 раз. Корневые системы ксерофитов часто экстенсивного типа, т. е. растения имеют длинные корни, распространяющиеся в большом объеме почвы, но мало разветвленные. Проникая на большую глубину, такие корни позволяют, например, пустынным кустарникам использовать влагу глубоких почвенных горизонтов, а в отдельных случаях и грунтовых вод. У других видов, таких, как степные злаки, корневые системы интенсивного типа. Они охватывают небольшой объем почвы, но благодаря густому ветвлению максимально используют влагу (рис. 4.17).
Наземные органы ксерофитов отличаются своеобразными чертами, носящими отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, хорошо заметная по густоте сети жилок в листьях, подводящих воду к тканям (рис. 4.18).
Эта особенность облегчает ксерофитам осуществлять пополнение запасов влаги, расходуемой на транспирацию. Структурные приспособления защитного характера у ксерофитов, направленные на уменьшение расхода воды, можно свести к следующему:
1. Общее сокращение транспирирующей поверхности за счет мелких узких, сильно редуцированных листовых пластинок.
2. Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона.
3. Защита листьев от больших потерь влаги на транспирацию благодаря развитию мощных покровных тканей — толстостенного или многослойного эпидермиса, нередко несущего различные выросты и волоски, которые образуют густое «войлочное» опушение поверхности листа.
4. Усиленное развитие механической ткани, предупреждающее обвисание листовых пластинок при больших потерях воды.

Рис. 4.17. Различные типы корневых систем:
А — экстенсивный (верблюжья колючка);
Б — интенсивный (пшеница
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев, перечисленными выше, имеют своеобразный внешний облик (чертополох, степные и пустынные полыни, ковыли, саксаул и др.), за что получили название склерофитов. Склерофиты (от греч. «склерос» — твердый, жесткий) не накапливают в себе влагу, а испаряют ее в большом количестве, постоянно доставая из глубоких слоев почвы. Тело этих растений жестковатое, сухое, иногда одревесневшее, с большим количеством механической ткани. При продолжительном прекращении подачи воды может наблюдаться сбрасывание листьев или части побегов, что приводит к сокращению испарения. Многие из ксерофитов переносят засушливое время года в состоянии вынужденного покоя.

Рис. 4.18. Различие жилкования (А), размеров и числа устьиц (Б)
у ксерофитов и мезофитов (из А. П. Шенникова, 1950):
1 — пустынный ксерофит — Psoralea drupaceae;
2 — лесной мезофит — Paris quadrifolia
Другая группа ксерофитов обладает способностью накапливать в своих тканях большое количество воды и получила название «суккуленты» (от лат. «суккулентус» — сочный, жирный). Водозапасающие ткани у них могут быть развиты в стеблях или листьях, поэтому они подразделяются на стеблевые суккуленты (кактусы, молочаи) и листовые суккуленты (алоэ, агава, молодило). Тело суккулентов, как правило, покрыто толстым кутинизированным эпидермисом и восковым налетом. Устьиц на поверхности тела почти нет. А если есть, то они мелкие, располагаются в ямках и большую часть времени закрыты. Открываются только на ночь. Все это предельно сокращает транспирацию. Характерной особенностью суккулентов является высокая поглощающая способность. В период дождей отдельные виды поглощают в себя больш„юе количество воды. Накопленную влагу суккуленты в дальнейшем медленно расходуют. Суккуленты произрастают в районах с жарким сухим климатом. Там, где хотя бы изредка проходят дожди кратковременные, но обильные, ливневые.
В целом же различные формы адаптации к водному режиму у растений и животных, выработанные в процессе эволюции, отражены в табл. 4.9.
Таблица 4.9
Не нашли, что искали? Воспользуйтесь поиском: