ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Ефект дії малих доз іонізуючого випромінювання
При дії різних типів іонізуючого випромінювання на біологічні середовища відбувається передача енергії малими кінцевими порціями (квантами) під час окремих актів взаємодії з речовиною. Для іонізації опромінюваного біологічного середовища необхідна мінімальна величина енергії, що визначається середнім потенціалом іонізації (табл. 8.3).
Під час дії іонізуючого випромінювання насамперед іонізуються молекули води - найбільш поширеного середовища будь-якого біологічного об'єкта, які перетворюються у вільні радикали Н і ОН. Як зазначалося, вільні радикали мають високу хімічну активність і вступають в реакції безпосередньо з біологічними молекулами. Починається ланцюгова реакція, коли в хімічні взаємодії вступає дедалі більше молекул. Викликані іонізуючим випромінюванням біохімічні реакції призводять до змін у ділянках кліткових структур (ДНК, мембрани, хроматиди тощо). При невеликих дозах опромінення захисні механізми організму відновлюють пошкодження в ДНК, виводять з організму пошкоджені клітини, нейтралізують їх системою імунного захисту. При великих дозах опромінення захисна система організму не в змозі ліквідувати наслідки іонізації, що призводить до порушення нормальної життєдіяльності клітин, тканин, органів і організму в цілому.
Таблиця 8.3. Середній потенціал іонізації для різних середовищ

Клітина, в якій іонізуючим випромінюванням викликані онкогенні зміни, тривалий час може залишатись в стані спокою. Для прояви її змінених властивостей необхідна дія на клітину активаторів (промоторів), які викличуть її прискорений поділ. В організмі всього налічується близько  клітин. Завжди є в наявності певна кількість ініційованих клітин, із яких тільки одиниці утворюють клани злоякісних клітин. Із збільшенням числа опромінень зростає кількість ініційованих клітин. Повторна радіаційна дія для цих клітин може служити промотором. Це пояснює кумулятивну дію іонізуючого випромінювання на біологічний об'єкт з погляду ризику віддалених ймовірних (стохастичних) наслідків.
клітин. Завжди є в наявності певна кількість ініційованих клітин, із яких тільки одиниці утворюють клани злоякісних клітин. Із збільшенням числа опромінень зростає кількість ініційованих клітин. Повторна радіаційна дія для цих клітин може служити промотором. Це пояснює кумулятивну дію іонізуючого випромінювання на біологічний об'єкт з погляду ризику віддалених ймовірних (стохастичних) наслідків.
Викликані зміни в генетичному апараті соматичних клітин можуть призвести до переродження клітин в злоякісні. Зміни в генетичному апараті статевих клітин в осіб діто-народжуваного періоду життя можуть спонукати до спадкових змін у нащадків опромінених.
Ведучими міжнародними організаціями (Міжнародна комісія з радіаційного захисту - МКРЗ, Міжнародне агентство з атомної енергії - МЛГАТЕ, Науковий комітет з дії атомної радіації при ООН - НКДАР ООН) приймається модель безпорогової лінійної залежності між дозою опромінення і її ефектом. Це передбачає, що навіть мінімальна радіаційна дія має біологічний ефект, зокрема радіаційний канцерогенез. Такий підхід базується на ймовірнісній (стохастичній) природі канцерогенезу. За оцінками ризик смертності від раку, викликаного радіаційним фоном порядку 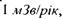 відповідно лінійній концепції складає близько 10 випадків на 1 млн. чоловік за рік. Це підтверджується даними, наведеними в табл. 8.2. До питання про лінійність залежності "доза-ефект" ми ще повернемося нижче.
відповідно лінійній концепції складає близько 10 випадків на 1 млн. чоловік за рік. Це підтверджується даними, наведеними в табл. 8.2. До питання про лінійність залежності "доза-ефект" ми ще повернемося нижче.
На думку багатьох дослідників, найбільш чутливою до іонізуючого випромінювання структурою клітини є ДНК, в якій іонізація може викликати розрив зв'язку між атомами. Одиничні і подвійні розриви викликають хромосомні спотворення (аберації). Поява хромосомних аберацій передує злоякісним пухлинам. Наявність хромосомних аберацій в лімфоцитах периферійної крові є ознакою передлейкозного стану, відповідає початковому етапу реакції на променеве навантаження.
Говорячи про ефект малих доз іонізуючого випромінювання, слід брати до уваги ще один дуже важливий фактор - тривалість їх дії. Справді, одну і ту ж саму, скажімо, поглинену (або еквівалентну) дозу можна отримати при відносно малих потужностях доз за рахунок збільшення часу їх взаємодії з речовиною (біооб'єктом, зокрема). Сказане підтверджується формулою
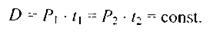 (8.39)
(8.39)
Звідси випливає досить очевидний факт: одне І те ж саме дозове навантаження іонізаційного випромінювання спостерігається для випадків, коли відношення потужностей доз обернено пропорційне часовим інтервалам дії цих потужностей, тобто
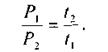 (8.40)
(8.40)
Приклад. Нехай потужність еквівалентної дози є досить малою: 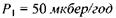 (приблизно тільки втричі більшою
(приблизно тільки втричі більшою
за природне фонове значення). Тоді еквівалентна доза цього випромінювання буде такою ж, як і випромінювання з суттєво більшою потужністю еквівалентної дози 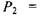 =
= 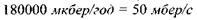 (потужність
(потужність 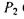 більш ніж в
більш ніж в
10 тисяч разів перевищує природне значення) за умови, що тривалість дії випромінювання малої потужності  складає 10 годин, тоді як тривалість дії випромінювання великої потужності
складає 10 годин, тоді як тривалість дії випромінювання великої потужності 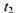 складає лише 10 с. Для даного прикладу маємо
складає лише 10 с. Для даного прикладу маємо  так що для обох випадків добуток потужності еквівалентної дози на час, тобто сумарна еквівалентна доза є величина стала:
так що для обох випадків добуток потужності еквівалентної дози на час, тобто сумарна еквівалентна доза є величина стала:
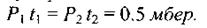
Ця ситуація значною мірою еквівалентна згаданій вище у параграфі 8.4.3 ситуації з популяційною дозою іонізуючого випромінювання, а саме: медико-біологічні ефекти (зокрема, кількість ракових захворювань) залишається сталою при одночасному пропорційному збільшенні еквівалентної дози 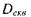 та зменшенні чисельності
та зменшенні чисельності  людської популяції, що опромінюється
людської популяції, що опромінюється 
І останнє зауваження щодо дії малих доз випромінювання на біооб'єкти. Воно стосується досить спірного і не до кінця з'ясованого явища підсилення дії фізико-хімічних (та, мабуть, і медико-біологічних) факторів при прямуванні параметрів, як: характеризують цей фактор (наприклад, концентрації речовини) до нуля. Іншими словами, йдеться про те, що не можна просто екстраполювати результати, які отримані при великих значеннях параметрів (концентрації, доз і т.п.) на малі значення цих параметрів, тобто сподівання на прямо пропорційне зменшення біологічного ефекту при зменшенні поглиненої дози не є апріорі вірним.
З математичної точки зору ефект підсилення часто описується такою формулою:
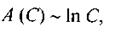 (8.41)
(8.41)
де  - певна властивість (фактор);
- певна властивість (фактор); 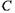 - відповідна змінна (параметр). Прикладом може бути наведена в параграфі 4.4.2 залежність осмотичного внеску до хімічного (електрохімічного) потенціалу від концентрації
- відповідна змінна (параметр). Прикладом може бути наведена в параграфі 4.4.2 залежність осмотичного внеску до хімічного (електрохімічного) потенціалу від концентрації  що подається виразом
що подається виразом
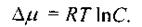 (8.42)
(8.42)
З формул (8.41) і (8.42) безпосередньо випливає, що коли концентрація С прямує до нуля, то величина \А\ або 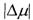 прямують до нескінченності. Більш уважний аналіз досить тонких математичних нюансів отримання подібних логарифмічних особливостей
прямують до нескінченності. Більш уважний аналіз досить тонких математичних нюансів отримання подібних логарифмічних особливостей 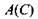 або
або 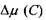 показує, що просто так прямувати змінну
показує, що просто так прямувати змінну 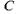 до нуля не можна (через використання так званої формули Стірлінга, яка перестає бути вірною при
до нуля не можна (через використання так званої формули Стірлінга, яка перестає бути вірною при 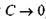 ). Водночас тенденція до збільшення фактора А при зменшенні параметра
). Водночас тенденція до збільшення фактора А при зменшенні параметра 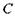 (хай не до нуля) залишається.
(хай не до нуля) залишається.
Подібні ефекти спостерігаються в фізико-хімічних дослідженнях деяких розчинів. Мабуть, саме вони лежать в основі дії гомеопатичних препаратів, а також в дії іонізуючого випромінювання в малих дозах на біологічні об'єкти. Звичайно, вкрай необхідне з'ясування особливостей молекулярних механізмів дії малих доз випромінювання, що потребує подальших теоретичних і експериментальних досліджень з боку фахівців різних галузей знань.
Не нашли, что искали? Воспользуйтесь поиском: