ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Форменные элементы крови
Эритроциты. Количество эритроцитов в крови различных животных не одинаково. Эритроциты отдельных видов животных отличаются между собой размерами, а также отсутствием (млекопитающие) или присутствием (птицы, земноводные) в них ядра.
Обладая большой удельной поверхностью, эритроциты могут адсорбировать на себе многочисленные органические и минеральные вещества, в том числе и газы, и транспортировать их к тканям. Основная их функция — дыхательная — неразрывно связана со свойствами содержащегося в них белка гемоглобина. Ниже приводится количество эритроцитов и их удельная поверхность в крови разных животных.
Вид животных
Лошади
Коровы
Овцы
Свиньи
Количество эритроцитов,
млн/мм
6—10
5,5—10
8—16
5,5—9
Удельная поверхность
эритроцитов, см2
17 000
16 000
19 100
15 900
Плотный остаток эритроцитов составляет около 40 % (35— 43 %) их массы. В основном этот остаток представлен гемоглобином (около 90%). У различных сельскохозяйственных животных абсолютное содержание гемоглобина в крови колеблется от 10 до 15 %, а в расчете на килограмм сухой массы телав среднем 12,7%. Кроме гемоглобина в составе эритроцитов имеются белки стромы, фосфолипиды, стериды, гликопротеиды и ряд ферментов — каталаза, карбоангидраза, холинэстераза, ферменты гликолиза.
Белки стромы, стериды, фосфолипиды и гликопротеиды образуют оболочку и внутреннюю структуру эритроцитов.
Ферменты эритроцитов катализируют реакции, определяющие специфику функций этих клеток. Так, каталаза с пособствует расщеплению перекиси водорода на воду и молекулярный кислород, устраняя появление в эритроцитах атомарного кислорода. Карбоангидраза обеспечивает синтез угольной кислоты из двуокиси углерода и воды в тканевых капиллярах, а в легочных альвеолах — разложение угольной кислоты на исходные продукты. Следовательно, этот фермент ответствен за связывание углекислоты в тканях и удаление ее из крови в альвеолах легких. Ацетилхолинэстераза участвует в регуляции проницаемости эритроцитов — установлена прямая связь между скоростью гемолиза эритроцитов и активностью холинэстеразы в них.
Эритроциты не обладают способностью к образованию внутриклеточных веществ, однако необходимость сохранения структуры клетки и гемоглобина, а также градиента концентрации ионов натрия и калия внутри эритроцитов и плазме требует постоянного притока энергии. Такую энергию эритроциты получают за счет происходящего в них гликолиза и частично при прямом окислении глюкозы.
- Из минеральных веществ в эритроцитах содержатся калий, натрий, кальций, магний, медь, железо, цинк и др., причем распределение этих элементов между плазмой и эритроцитами не одинаковое. Следует отметить, что цинк в эритроцитах почти полностью содержится в составе карбоангидразы, железо — в составе гемоглобина.
Выполнение эритроцитами дыхательной функции обусловлено наличием в их составе белка гемоглобина. В легких гемоглобин присоединяет к себе кислород и транспортирует его к тканям. В тканях, отдав им кислород, эритроциты связывают углекислый газ (около 90% от образующегося) и транспортируют его к легким.
Гемоглобин относится к хромопротеидам и состоит из протеина— глобина (96% молекулярной массы) и простетической группы — тема (4 %).
Гемоглобин разных животных отличается строением своей белковой части — глобина и всегда имеет одинаковую простетическую группу — гем. Молекулярная масса гемоглобина составляет 64000—68000, он хорошо кристаллизуется. Кристаллы гемоглобина вишнево-красного цвета, оксигемоглобина - ярко-красного. Изоэлектрическая точка гемоглобина находится при рН = 6,8, а для оксигемоглобина — при рН = 6,6—6,7. В водных растворах гемоглобин быстро разрушается.
В крови эмбрионов содержится эмбриональный или примитивный гемоглобин, который постепенно замещается на гемоглобин F (лат. «Fetus»—плод). У взрослых особей в крови содержится гемоглобин А (лат. «Adultus»—взрослый). Гемоглобин А представлен тремя фракциями: Аь А2 и А3. Отличаются они между собой набором и последовательностью аминокислот в полипептидных цепях.
При ряде заболеваний в крови могут появиться и другие виды гемоглобина — S, С, D, Е и т. д. Они возникают в результате изменения последовательности расположения аминокислот в полипептидных цепях. Положение аминокислот в р-полипептидной цепи разных видов гемоглобина следующее:
Гемоглобин А (норма) Гемоглобин S (болезнь) Гемоглобин С (болезнь) Гемоглобин D (болезнь)
6 7
Глю — Глю Вал — Глю Лиз — Глю Глю — Глю
Замена даже одной аминокислоты (в положении 6 или 7) влечет за собой изменение физико-химических свойств гемоглобина и частичную утрату его способности к связыванию кислорода.
Так, при серповидной анемии в крови больных животных появляется гемоглобин S, растворимость которого значительно ниже, чем у гемоглобина А. Это приводит к тому, что отдельные молекулы гемоглобина соединяются между собой в межмолекулярные агрегаты с образованием осадка и последующей деформацией эритроцитов (последние приобретают серповидную форму). Серповидная анемия сопровождается нарушением проницаемости эритроцитарной мембраны для фосфатов и фосфорных эфнров глюкозы. Такие изменения способствуют снижению устойчивости эритроцитов к гемолизу и ускорению их разрушения.^
Свободный гем с кислородом образует прочное соединение, но, находясь в составе гемоглобина, он приобретает новые свойства.
В основе структуры гема лежит протопорфириновое кольцо, в центре которого расположен атом двухвалентного железа. Железо имеет шесть координационных связей. Четыре из них (две главные и две побочные) заполнены лигандными группами протопорфирина, в результате чего образуется плоский квадратный комплекс, который и получил название тема. Остальные две координационные связи железа направлены перпендикулярно плоскости молекулы тема. Одной из этих связей гем соединяется с глобином, а шестая остается свободной (дезоксигемоглобин) или обратимо замещается кислородом (оксигемоглобин) или другими лигандами, например окисью углерода.
Гем, соединяясь с глобином, располагается в углублении, образованном третичной структурой последнего. Полипептидиые цепи а-, р-функциональных субъединиц глобина в дезоксигемоглобине соединены попарно, имеется четыре мостика между а-цепями и два — между р-цепями. В оксигемоглобине указанные связи отсутствуют, что повышает его сродство к кислороду и ускоряет процесс оксигенации (присоединение кислорода).
Указанные особенности строения и свойство гемоглобина обеспечивают ему кооперативное связывание четырех молекул кислорода: каждый гем присоединяет кислород не самостоятельно, независимо друг от друга, а совместно, кооперативно. Кооперативное состоит в том, что каждую последующую молекулу кислорода гемоглобин присоединяет (и отдает) легче, чем предыдущую. Это объясняется тем, что после присоединения первой молекулы кислорода происходит изменение конформации одной из полипептидных цепей глобина и связанный с ней гем приобретает повышенное сродство к кислороду. Изменение первой полипептидной цепи автоматически передается на вторую полипептидную цепь, а от первой функциональной субъединицы гемоглобина конформационные изменения передаются на вторую, в результате чего последовательно происходит повышение сродства всех гемов гемоблобина к кислороду. Если принять константу взаимодействия первого тема с кислородом за единицу, то скорость присоединения кислорода к последующим темам выразится соотношением 1: 4: 24: 9.
Оксигемоглобин — основное кислородное производное гемоглобина, легко диссоциирующее на гемоглобин и О2. Скорость прямой и обратной реакции при взаимодействии гемоглобина и кислорода зависит от величины парциального давления кислорода в воздухе, с которым контактирует кровь, находясь в капиллярах легочных альвеол.
Гемоглобин крови животных и человека связывает разные количества кислорода даже при максимальном давлении последнего. Так, 1 г гемоглобина человека связывает 1,34 мл кислорода, лошади—1,33, коровы—1,0—1,5, свиньи—1,68, собаки—1,55—1,65 мл. По количеству кислорода, связываемого гемоглобином, содержащимся в 100 мл крови, определяют кислородную емкость крови. Для разных видов животных она колеблется в пределах от 10,1 до 19,5 мл. Оксигенация гемоглобина происходит быстрее, чем распад оксигемоглобина:
НЬ+02->НЬО2 за 0,001 с НЬО2->-НЬ + О2 за 0,05 с /
Распад оксигемоглобина на гемоглобин и кислород усиливается при понижении атмосферного давления, повышении температуры, пропускании через кровь азота, углекислоты и в ряде других случаев.
В артериальной крови практически весь гемоглобин находится в виде оксигемоглобина. Это обеспечивается относительно высоким парциальным давлением кислорода в легких, через которые проходит кровь. Во вдыхаемом воздухе 20,9 % кислорода и только 0,03 % углекислоты, тогда как в выдыхаемом воздухе 15,5—16,3% кислорода (на 22,1—26,3% меньше, чем во вдыхаемом) и 4,1 % углекислого газа (в 136 раз больше, чем в атмосфере). Учитывая то обстоятельство, что толщина эпителиальной стенки, отделяющей кровь от воздуха в альвеолах, составляет 0,004 мм, а поверхность крови в легких крупных животных превышает площадь в 90—100 м2, создаются благоприятные условия для диффузии кислорода в кровь, а двуокиси углерода в воздух. В процессе дыхания организм использует около 5 % кислорода, содержащегося в воздухе. Процесс присоединения молекулярного кислорода к гемоглобину не сопровождается изменением валентности железа тема, поэтому он получил название оксигенация, а не окисление. Образующийся при оксигенации оксигемоглобин в тканях распадается на гемоглобин и кислород, после чего кислород усваивается тканями.
Образование оксигемоглобина зависит от парциального давления кислорода в воздухе. При уменьшении этого давления количество образующегося оксигемоглобина уменьшается -возникает состояние гипоксемии. Недостаток кислорода в крови вызывает его недостаток и в тканях. Такое явление называется гипоксией. При наличии во вдыхаемом воздухе 20,9% кислорода около 99 % гемоглобина крови превращается в оксигемоглобин, при 10 % кислорода около 90 % гемоглобина превращается в оксигемоглобин, а при 2,5 % —только 50 %, это свидетельствует о существовании адаптационных механизмов в организме человека и животных к изменению концентрации кислорода в воздухе. В основе адаптационного механизма, немаловажную роль играет кооперативность при взаимодействии гемоглобина с кислородом.
Аноксемия возникает при восхождении на высокие горы (горная болезнь), при вдыхании инертных газов, а также при нарушении нормальной функции легких (воспалительные процессы, отеки, эмфизема).
Гемоглобин участвует и в переносе CO2J В межтканевых жидкостях парциальное давление СО2 около 8,0 кПа (60 мм рт. ст.), а в венозной крови — около 6 кПа (47 мм рт. ст.). Эта разница обеспечивает диффузию СО2 из тканей в венозные сосуды./В крови около 20 % СО2 соединяется с гемоглобином карбамидными связями ро месту свободных аминогрупп глобина (образуется карбогёмоглобин)! и около 80 % СО2 превращается в гидрокарбонаты калия. В легких гидрокарбонаты разрушаются до угольной кислоты, которая распадается на воду и углекислый газ и эти продукты удаляются из организма с выдыхаемым воздухом. Образование угольной кислоты в тканях и ее распад в легких до СО2 и Н2О осуществляется ферментом карбоангидразой.
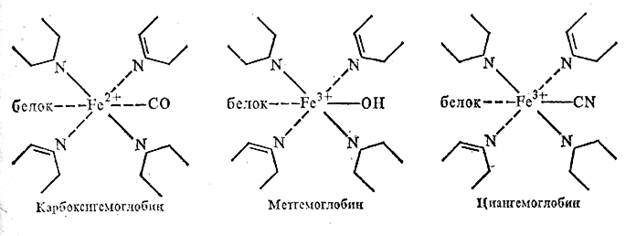
Гемоглобин помимо оксигемоглобина и карбогемоглобина может образовать и ряд других производных. К этим производным относятся метгемоглобин, карбоксигемоглобин, циангемоглобин. Образование метгемоглобина (НЬ—ОН) происходит при попадании в кровь различных окислителей (метиленовая синь, красная кровяная соль, мышьяковистый водород, бензол, нитробензол, анилин). В присутствии различных окислителей железо тема окисляется до трехвалентного состояния и прочно соединяется с группой —ОН, утрачивая способность присоединять кислород. Под действием дегидрогеназ тканей метгемоглобин постепенно восстанавливается до гемоглобина. Однако превращение больших количеств гемоглобина крови в метгемоглобин (50 % и больше) вызывает гипоксию и может послужить причиной смерти.
/Гемоглобин очень активно соединяется с угарным газом, образуя при этом карбоксигемоглобин (НЬ-—СО). Сродство гемоглобина к окиси углерода примерно в 300 раз выше, чем к кислороду, в связи с чем при наличии в воздухе даже 0,1 % ионов углерода больше половины гемоглобина превращается в карбоксигемоглобин^] Карбоксигемоглобин, как и метгемоглобин, не обладает способностью к оксигенации, поэтому образование больших его количеств приводит к гипоксии тканей, и смерти животного., В карбоксигемоглобине железо остается
двухвалентным. Находясь в крови животных, отравленных угарным газом, НЬ—СО придает ей алый цвет. В отличие от артериальной крови, содержащей НЬ—О2, она не темнеет при попадании в среду с низким парциальным давлением кислорода, т. е. карбоксигемоглобин не переходит в гемоглобин.
Циангемоглобин (Hb—CN) возникает при попадании в организм синильной кислоты и ее солей. В циангемоглобине группа —CN присоединяется к железу тема довольно прочно (железо становится трехвалентным элементом):
Гемоглобин и его производные дают характерные спектры поглощения, на основании чего их можно идентифицировать.
Распад гемоглобина. Эритроциты в организме, как и все клетки, функционируют в течение определенного периода времени (у млекопитающих от 30 до 120 дней), после чего происходит их распад. Местом распада эритроцитов являются селезенка, печень, костный мозг. Освобождающийся при их разрушении гемоглобин после ряда последовательных изменений распадается до свободных аминокислот (глобин), а входящий в его состав гем постепенно превращается в желчные пигменты—билирубин и биливердин, которые конъюгируют с глюкуроновой кислотой и в дальнейшем удаляются из организма частично с калом — стеркобилин и частично с мочой— уробилин (подробно см. «Обмен хромопротеидов»),
Лейкоциты. Над выпавшими в осадок эритроцитами образуется небольшой слой лейкоцитов — белых клеток крови. Лейкоциты играют очень важную роль в защите организма. Они могут захватывать и переваривать бактерии и другие инородные тела, попавшие в организм.
В отличие от эритроцитов лейкоциты имеют ядро и все другие органеллы, присущие соматическим клеткам. Наличие ядра и рибосом свидетельствует о том, что в лейкоцитах постоянно синтезируются различные белки. В числе последних множество ферментов, среди которых наиболее активны оксидоредуктазы и гидролазы. Высокая активность гидролитических ферментов — катепсинов, липаз, гликозидаз, амидаз — обеспечивает.участие, ле.йконитов в пеоеваоиванин чужеродных органических веществ. Лейкоциты имеют активную систему аэробных дегидрогеназ и других ферментов дыхательной цепи. Освобождающаяся при окислении органических веществ в лейкоцитах энергия используется также для удержания структуры клеток, осуществления активного транспорта веществ через их мембрану против градиента концентрации.
Лейкоциты могут обволакивать эластической мембраной чужеродные тела и клетки и в последующем переваривать их. Это явление названо фагоцитозом. Он сопровождается резким увеличением поглощения кислорода и глюкозы лейкоцитами, повышением распада гликогена в них, нарастанием активности ферментов. Например, активность ферментов пентозного цикла при фагоцитозе увеличивается в 15 раз. Фагоцитоз также сопровождается образованием большого количества Н2О2, концентрация которой достигает такой величины, что обнаруживается ее бактерицидное действие.
В лейкоцитах в больших количествах вырабатывается гистамин. Эти клетки активно участвуют в образовании целого ряда иммуноглобулинов; Особенно высоко активны в этом отношении такие разновидности лейкоцитов, как плазмоциты.
Лейкоциты вырабатывают специфический фермент, напоминающий по своему действию лизоцим. Назначение этого фермента состоит в том, что он разрушает полисахаридные комплексы бактериальных клеток и тем самым способствует ускорению их фагоцитоза лейкоцитами.
Тромбоциты. Кровяные пластинки, или тромбоциты, играют важную роль в образовании кровяного сгустка при свертывании крови, его ретракции и сужении сосудов. На их поверхности сорбируются плазматические факторы коагуляции и другие биологически активные вещества (например, гистамин, серотонин), способствующие свертыванию крови и уменьшению кровотечения за счет сужения просвета кровеносных сосудов.
Химический состав и особенности обмена тромбоцитов изучены еще недостаточно, хотя некоторые биохимические особенности этих клеток известны. Так, в отличие от эритроцитов и плазмы в них много фосфатов и совсем мало гидрокарбонатов. Тромбоциты богаты фосфатидами и кальцием, натрий в них находится в связанном с фосфатидами состоянии. Участие тромбоцитов в свертывании крови зависит от наличия в их составе ненасыщенных жирных кислот — чем их больше в тромбоцитах, тем активнее происходит образование кровяного сгустка. Непредельные жирные кислоты входят в состав специфического липопротеида, выполняющего роль одного из факторов свертывания крови.
Тромбоциты содержат в себе фактор, нейтрализующий антикоагуляционное действие гепарина. Действие тромбоцитов на ретракцию кровяного сгустка основано на наличии в их составе молекул, напоминающих по строению и свойствам актомиозин мышц и способных сокращаться при наличии АТФ. Содержание АТФ в тромбоцитах достаточно высокое — в 100—150 раз больше, чем в эритроцитах.
Лимфа
Лимфа — прозрачная жидкость, заполняющая просвет лимфатических сосудов, межтканевых и межклеточных пространств. По составу и физико-химическим свойствам она во многом напоминает плазму крови, отличаясь от нее более низким содержанием белков. Плотность лимфы 1,012—1,026, относительная вязкость 1,0—1,5; поверхностное натяжение 1,00—1,02, депрессия 0,58—0,59, рН 7,6—7,7.
Количество лимфы у крупного рогатого скота достигает 95—100 л, у лошадей —70—80, у собаки —0,6 л.
Лимфа, полученная из разных участков тела, отличается между собой. Так, в лимфе нижних конечностей содержание белков не превышает 3 %, в лимфе кишечника их 5—6 %, а в лимфе печени 6—8 %.- К белкам лимфы относят альбумин, глобулины, фибриноген, протромбин. Соотношение между альбуминами и глобулинами в лимфе крупного рогатого скота следующее (%): альбуминов — до 45, а-глобулинов —до 20, γ-глобулинов— до 12, у-глобУлннов — Д° 23. Фибриногена в лимфе 0,08—0,10, т. е. в 4—5 раз меньше, чем в плазме.
В лимфе всегда содержатся мочевина, аминокислоты, креатинин, азотистые основания, жиры, холестерол, фосфатиды, сахар, хлориды, фосфаты, натрий, калий, кальций, магний, ферменты, гормоны, витамины и другие элементы и вещества. Содержание перечисленных веществ в лимфе зависит от того, когда взята лимфа для исследований — до или после кормления. У животных до кормления лимфа содержит очень мало жиров (до 0,01 %), а после кормления их количество может достигать 3 % и больше. Количество сахара в лимфе относительно постоянное и для разных животных колеблется от 0,05 до 0,10%.
Из форменных элементов в лимфе присутствуют в небольшом количестве лимфоциты.
Не нашли, что искали? Воспользуйтесь поиском: