ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Геохронологическое подразделение истории Земли
| Эра | Период | Эпоха | Продолжи- тельность (в млн. лет) | Время от начала периода до наших дней (в млн. лет) | Преобладающие формы жизни | Климат | Геологические условия |
| Кайнозой (новая) | Четвертичный | Совре- менная | 0,02 | Травянистые растения, млекопитающие. Эпоха человека | теплый | Конец последнего ледникового периода | |
| Поейсто-цен | 1,7 | Вымирание многих видов растений и крупных млекопитающих. Появление человека | Холодный | Повторные оледенения (четыре ледниковых периода) | |||
| Третичный | Плиоцен | Цветковые растения. Млекопитающие (появления лонов, лошадей, верблюдов, сходных с современными) | Прохладный | Поднятие отдельных гор. Остепенение суши. Вулканическая активность | |||
| Миоцен | Появление первых человекообразных обезьян | Прохладный | Образование гор. Вулканическая активность | ||||
| Олигоцен | Леса. Однодольные цветковые растения. Предшественники многих современных родов млекопитающих (включая человекообразных обезьян и людей) | теплый | Материки низменные | ||||
| Эоцен | Пацентарные млекопитающие | теплый | Горы размыты. Внутриконтинентальных морей нет | ||||
| Палеоцен | Примитивные млекопитающие. Угасание царства динозавров | ||||||
| Мезозой | Мел | Покрытосесенные однодольные и двудолные растения. Первые цветки. Динозавры. Примитивные млекопитающие | Образуются горы, отложения мела. Сланцев | ||||
| Юра | Цикадофиты и хвойные. Динозавры. Древнейшие птицы, насекомоядные, сумчатые | Материки довольно возвышены. Мелководные моря | |||||
| Триас | Голосеменные, динозавры | Теплый, усиливается засушливость | Материки приподняты над уровнем моря |
Продолжение табл.32.
| Эра | Период | Эпоха | Продолжи- тельность (в млн. лет) | Время от начала периода до наших дней (в млн. лет) | Преобладающие формы жизни | Климат | Геологические условия |
| Герцинское горообразование | |||||||
| Палеозой (древний) | Пермь | Голосеменные, насекомые, пресмыкающиеся. Появление предков динозавров | Аридный | Материки приподняты, периодическое оледенение | |||
| Карбон (каменноугольный) | Папоротникообразные, плауны, голосеменные, первые настоящие семена. Земноводные | Теплый, влажный | Болота, поднятие суши, материки низменнные | ||||
| Девон | Первые леса (папоротники). Первые настоящие листья. Рыбы. | Аридный | Небольшие внутриконтинентальные моря. Оледенение | ||||
| Силур | Водоросли, хвощи, плауны. Морские паукообразные (скорпионы). Начался выход животных на сушу | Засушливый | Обширные внутриконтинентальные моря. Поднятие суши | ||||
| Ордовик | Морские водоросли. Наутилоидные. Трилобиты, моллюски | Теплый, включая Арктику | Заметное погружение суши | ||||
| Кембрий | Морские водоросли, «взрыв» морских многоклеточныех. Трилобиты и плеченогие, губки и иглокожие | Умеренный | Материки низменные | ||||
| Второе великое горообразование | |||||||
| Протерозой | Примитивные водоросли и водные грибы. Морские простейшие. В конце – моллюски, черви и др. Морские беспозвоночные | Интенсивное осадкообразование. Вулканическая активность, обширная эрозхия и многократные обледенения | |||||
| Первое великое горообразование | |||||||
| Архей | Ископаемые не обнаружены (обнаружено органическое вещество в скальных породах) | Значительная вулканическая активность. Слабое осадкообразование. Обширная эрозия |
Копытные в ходе эволюции разошлись на группу с четным числом пальцев (коровы, овцы, верблюды, олени, жирафы, свиньи и гиппопотамы) и группу с нечетным числом пальцев (лошади, зебры, тапиры, носороги). В эоцене существовали китообразные формы, давшие начало китам и дельфинам. В олигоцене широко были распространены также предковые формы человекообразных обезьян и человека.
В миоцене и плиоцене древесная растительность вытесняется травянистыми формами. Наступает расцвет покрытосеменных растений, членистоногих и млекопитающих. Конец третичного периода датируется появлением всех современных семейств животных и растений.
Следующий период (четвертичный) этой эры, длящийся последние 1,5-1,7 млн лет, состоит из плейстоценовой и современной эпох. Для плейстоцена характерны четыре последовательных оледенения. Растения и животные, которые были распространены в плейстоцене, сходны с современными растениями и животными. Однако в плейстоцене вымерли многие растения (особенно лесные) и млекопитающие (мамонты, саблезубые тигры). Появились многие травянистые формы растений, но, самое главное, появился человек. Современная эпоха началась около 11 000 лет назад, когда отступил последний ледник.
Хотя в палеонтологической летописи существуют многие пробелы, тем не менее она полностью раскрывает происхождение многих существующих в настоящее время видов животных и растений. Например, очень хорошо прослежено происхождение лошадей, слонов, парнокопытных, хищников.
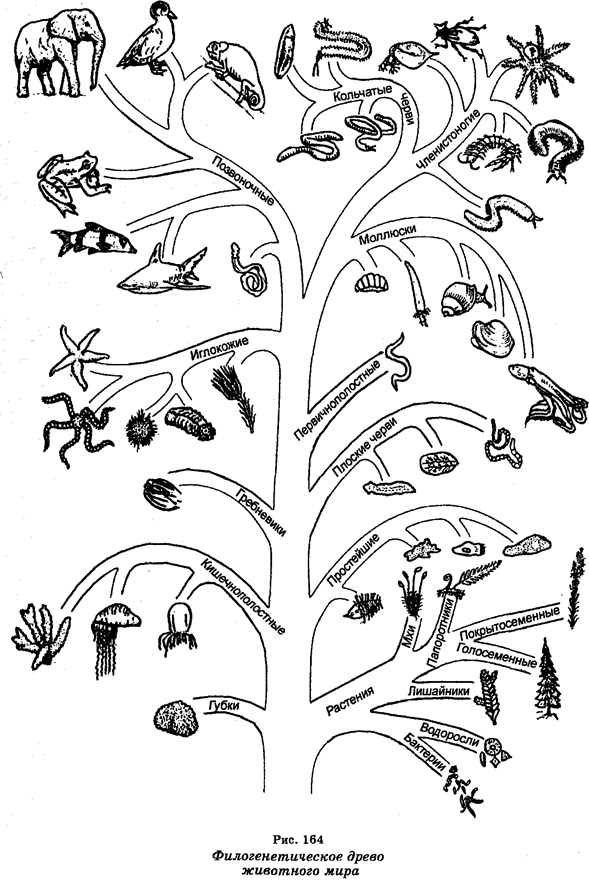
Эволюцию на уровне крупных таксонов называют макроэволюцией. Основополагающие представления о макроэволюции в целом сводятся к тому, что все современные живые существа являются измененными потомками одной или нескольких форм, существовавших ранее. Более молодые по происхождению формы животных и растений произошли от предков, менее отличавшихся друг от друга, а те, в свою очередь, развились от общего единого источника (рис. 164).
Считают, что первыми возникли архебактерии. Примерно 3,5 млрд лет назад. Затем возникли истинные бактерии, а от них ответвились вирусы, цианобактерии, красные водоросли и зеленые жгутиковые. От последних ответвились водоросли (от которых позднее произошли, в свою очередь, высшие растения) и бесцветные ризофлагелляты, из которых развились диатомовые водоросли, плесневые грибы, губки и простейшие. Часть простейших стала паразитической.
Вторым крупнейшим этапом эволюции была трансформация одноклеточных организмов в многоклеточные. Считают, что первые эукариоты появились 1,5 млрд лет назад. Их появление есть крупнейший эволюционный переход, ибо в основе появления эукарио-тов лежит формирование митоза и развитие клеточных структур. Из цилиарных простейших (цилиофор) развились многоклеточные (синтициальные) плоские черви. Последние разветвились на пять линий, одна из которых повела к иглокожим и хордовым. Остальные линии повели к большинству других типов животного мира.
Первыми позвоночными были остракодермы, которые развились из беспозвоночных, а из остракодерм развились миноги и панцирные акулы. Хрящевые рыбы (акулы и скатовые) и костные рыбы развились из панцирных акул, а из костных рыб развились кроссоптериги, которые являлись примитивными рыбами и которые затем разошлись на две эволюционные ветви, одна из которых привела к дипловертебронам (ранним земноводным), называемым стегоцефалами. Дипловертеброны были предшественниками современных амфибий и пресмыкающихся. Первые пресмыкающиеся (котилозавры) разветвились на ветви, давшие начало динозаврам и морским пресмыкающимся, которые затем вымерли, а также черепахам, крокодилам, ящерицам, змеям, птицам и млекопитающим.
Эволюционная линия, ведущая к человеку, начинается от ту-пай и ведет к ранним приматам, разветвляясь затем на обезьян Нового света, обезьян Старого света, понгид и гоминид. От понгид (настоящих обезьян) произошли гориллы, орангутанги и шимпанзе, а от гоминид произошел непосредственный предшественник современного человека (см. гл. XVI).
Главные направления эволюции сформулированы А. Н. Север-цовым (1866-1936), который впервые четко определил различия между прогрессивным и регрессивным развитием, обосновал понятия о морфофизиологическом и биологическом прогрессе, а также о морфофизиологическом и биологической регрессе.
Морфофизиологический прогресс — это изменения структуры и общей жизнедеятельности на пути эволюции от простых форм к сложным, от одноклеточных организмов к многоклеточным. Напротив, Морфофизиологический регресс — это упрощение организации и жизнедеятельности, выражающееся в редукции органов (например, хорды у оболочников) на пути эволюции.
Следствием морофофизиологического прогресса является биологический прогресс. Он характерен для организмов, которые характеризуются возрастанием приспособленности к окружающей среде, что ведет к увеличению их численности и расширению их ареалов. Однако биологический прогресс может быть результатом не только морфофизиологического прогресса. Иногда от является и результатом морфофизиологического регресса
В конечном итоге биологический прогресс ведет к процветанию вида.
В современную эпоху процветающими являются покрытосеменные растения, насекомые, костные рыбы, птицы, млекопитающие.
Значение биологического прогресса заключается в том, что он играет важную роль в эволюции и обеспечивает главные направления эволюции в виде ароморфозов, идиоадаптаций и общей дегенерации.
Ароморфозы — это крупные изменения организмов, которые поднимают на новый более высокий уровень их морфофизиологическую организацию и жизнедеятельность. Кроме того, они открывают организмам путь к проникновению в новую среду обитания. Ароморфозы возникают скачками. Примерами ароморфозов А. Н. Северцов называл развитие легких и четырехкамерного сердца, превращение парных плавников рыб в парные конечности земноводных. По А. Н. Се-верцову, ароморфозы обозначили возникновение организмов новых классов и типов организмов, завоевание организмами новых просторов. У млекопитающих ароморфозами были: появление живорождения, шерстного покрова, постоянной температуры тела и др.
Идиоадаптация — это изменения, которые не приводят к повышению уровня организации организмов, но сопровождаются приданием им большей приспособленности к существующим условиям жизни, больших возможностей прогрессивного развития. Примером идиоадаптаций являются приспособления насекомых к жизни в разных средах (в воде, почве, воздухе), к разным типам питания. Другим примером служит разнообразие форм у насекомоядных (землеройка, крот, еж, выхухоль и др.), что позволило занять им разные экологические ниши.
Общая дегенерация — это изменения, которые заключаются в снижении уровня организации живых существ и в снижении активности отдельных или ряда органов. Примерами общей дегенерации А. Н. Северцов называл биологию саккулины из группы паразитических ракообразных (крабов), а также биологию оболочников. Выживаемость этих организмов обеспечивается чрезвычайным разнообразием их личиночных форм.
Следствием морфофизиологического регресса является биологический регресс. Биологический регресс — это снижение приспособленности организмов к среде, сокращение их численности и ареалов. Конечным итогом биологического регресса является вымирание организмов.
Учение А. Н. Северцова о главных направлениях эволюционного процесса было развито дальше в трудах И. И. Шмальгаузена, который рассматривал организм как целое в историческом и индивидуальной развитии. Среди главных направлений эволюционного процесса И. И. Шмальгаузен большое внимание уделял различным формам ароморфозов.
Доказательства эволюции получены в разных науках. Классические доказательства эволюции получены, прежде всего, в палеонтологии в результате изучения ископаемых организмов, живших в прошлые эпохи. Предполагают, что в ходе эволюции вымерло около 200 000 видов животных. В более глубоких слоях Земли обнаруживаются остатки более древних форм жизни, тогда как в поверхностных слоях находят остатки более поздних форм. Можно сказать, что история жизни на Земле написана на языке ископаемых остатков. Палеонтологический материал дает также основания судить о темпах и направлениях эволюции.
Доказательства эволюции получены в биогеографии, которая является наукой о распространении растений и животных. В биогеографии различают шесть биогеографических областей. Каждая из этих областей характеризуется специфическими обитателями (растениями и животными), называемыми эндемиками, под которыми понимают организмы видов, родов и таксонов, ограниченных в своем распространении определенными территориями.
В Палеоарктической области (Европа, африканский север от Са-хары, часть Азии к северу от Гималаев, Азорские острова и острова Зеленого мыса) эндемичными, если говорить о животных, являются кроты, олени, быки, овцы, козы, скворцы и сороки.
В Неоарктической области (Гренландия и Северная Америка) эндемичными являются горные козлы, луговые собачки, опоссумы, скунсы, еноты, сойки и американские грифы. Кроме того здесь встречаются формы, андемичные для палеоарктической области.
В Неотропической области (Южная и центральная Америка, юг Мексики и острова Вест-Индии) обитают альпаки, ламы, цепко-хвостые обезьяны, тапиры, ленивцы, вампиры, муравьеды и многие виды птиц, не встречающиеся в других частях земного Шара.
В Эфиопской области (Африка к югу от Сахары, Мадагаскар) обитают шимпанзе, гориллы, зебры, носороги, трубкозубы, бегемоты, жирафы, многие виды птиц, пресмыкающихся и рыб, не обнаруживаемые в других областях.
В Восточной области (Индия, Цейлон, Индокитай, юг Китая, Малайский полуостров и отдельные острова Малайского архипелага) обитают орангутанги, черные пантеры, индийские слоны, гиббоны и долгопяты. В Австралийской области (Австралия, Новая Зеландия, Новая Гвинея и др. острова Малайского архипелага) эндемичными являются утконосы, кенгуру, вомбаты, коала и другие сумчатые животные. Эндемичными являются бескрылые птицы эму и казуар, а также птица-лира и какаду.
Географические закономерности, характерные для фауны, присущи и флоре этих биогеографических областей.
Одно из основных положений биогеографии заключается в том, что каждый вид растений и животных возникал только однажды и только в одном месте (центре происхождения), откуда он расселялся до тех пор, пока не встречал какую-нибудь преграду, например, географическую, климатическую, пищевую и т. д. Географические ареалы близких видов, как правило, не совпадают, но они и не очень отдалены один от другого.
Особенности географического распространения животных и растений являются отражением специфики эволюции каждого вида. Например, сходство фауны Неоарктической и Неотропической областей обусловлено относительно недавним геологическим разделением Северной Америки и Евразии в результате образования Берингова пролива. Наличие в Австралии однопроходных и сумчатых, которых нет ни в одной из других областей, является результатом того, что этот континент давно (в мезозое) был изолирован от других частей суши. Примитивным млекопитающим — аборигенам Австралии не довелось конкурировать с более приспособленными плацентарными млекопитающими, поэтому они дали начало формам, легко приспособившимся к местным условиям.
Далее, сейчас аллигаторы встречаются только в реках юго-востока США и в р. Янцзы (КНР), тогда как магнолии произрастают только в восточной части США, в КНР и Японии. В начале кайнозоя Северная Америка соединялась с Азией (на месте теперешнего Берингова пролива). При тогдашнем теплом климате аллигаторы и магнолии на этой территории были распространены повсеместно. В дальнейшем из-за поднятия Скалистых гор и похолодания (оледенения) аллигаторы и магнолии погибли повсеместно, кроме юго-восточной части США и восточной части КНР, где не было оледенения. Поскольку виды магнолий и аллигаторов были разделены в отдаленных частях земного шара на протяжении многих миллионов лет и эволюционировали обособленно, то в современную эпоху, оставаясь близкородственными видами, они несколько различаются между собой.
Еще Ч. Дарвин отмечал различия островных и материковых флор и фаун. Например, на Галапагосских островах, возраст которых составляет всего лишь 3—5 млн лет и которые никогда не были связаны с материком, отсутствуют лягушки и жабы, ибо ни они, ни их яйца не смогли бы выжить в морской воде, с которой неизбежно пришлось бы столкнуться мигрирующим на этот остров животным. На этих островах нет также наземных млекопитающих, но на одном из этих островов встречаются гигантские ящерицы и черепахи, которые очень близки некоторым формам, обитающим на побережье Эквадора. Это указывает на эволюцию растений и животных после заселения этих островов.
В систематике главное доказательство эволюции заключается в том, что все живые существа можно расположить в иерархическую систему таксономических единиц — виды, роды, семейства, отряды, классы и типы. Это означает, что все организмы связаны между собой филогенетически в результате существующих между ними эволюционных взаимоотношений. Принадлежность организмов к тем или иным систематическим группам свидетельствует о том, что большинство промежуточных форм, существовавших в прошлом, вымерло. Если бы виды всех существовавших в прошлом организмов жили до настоящего времени, то классифицировать живой мир на таксономические группы было бы невозможно. Будучи генетически различными, виды представляют собой независимо эволюционирующие и репродуктивно изолированные единицы. Поскольку можно предполагать, что у генетически сходных видов общий предок существовал в менее отдаленном прошлом по сравнению с генетически различными видами, то степень генетических различий является, по существу, мерой, на основе которой сейчас совершенствуют филогенетическое древо,
Степень генетических различий между видами определяют либо прямо путем изменений последовательностей нуклеотидов в генах, либо косвенно путем изменений последовательностей нуклеотидов в рРНК, или последовательностей аминокислот в белках. Результаты сравнения последовательностей ДНК разных организмов позволяют определить количество пар нуклеотидов, в которых в ходе эволюции имели место замены азотистых оснований (табл. 33), тогда как сравнение белков от разных организмов позволяет определить различия в аминокислотных последовательностях, т. е. судить о близости организмов (рис. 165) и о связи последовательностей со скоростью эволюции (табл. 34, рис. 166). На основе данных о филогении отдельных белков строят филогенетическое древо, которое, как показано для цитохрома С, совпадает с филогенетическим древом, построенным по ископаемым останкам. На основе реконструкции филогении и определения степени генетических различий по аминокислотным последовательностям ряда белков считают, что гены, кодирующие эти белки у животных, происходят от общего предка.
Таблица 33
Не нашли, что искали? Воспользуйтесь поиском: