ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Генетика поведения некоторых видов насекомых
МЕДОНОСНАЯ ПЧЕЛА
Медоносная пчела Apis mellifera – социальное насекомое, живущее большими семьями. Единственную плодовитую самку колонии – пчелиную матку – можно искусственно оплодотворить, используя сперму трутня нужного генотипа. Как известно, в размножении этих перепончатокрылых бесплодные рабочие особи колонии имеют 75% общих генов. В связи с таким способом размножения оценку генетической изменчивости пчел проводят, беря за единицу для анализа не отдельную особь, а пчелиную семью.
Классическая работа по генетике поведения пчел была проведена Н. Ротенбулером несколько десятилетий назад (RothenbuMer, 1967).
Пчеловодам было известно, что пчелы некоторых пород устойчивы к инфекции: не болеют американской гнильцой. Породы, устойчивые к этому заболеванию, имеют к своем поведенческом репертуаре специфические "гигиенические реакции". Пчелы каким-то образом обнаруживают ячейки, в которых находятся инфицированные личинки, и выбрасывают их из улья, препятствуя тем самым распространению болезни.
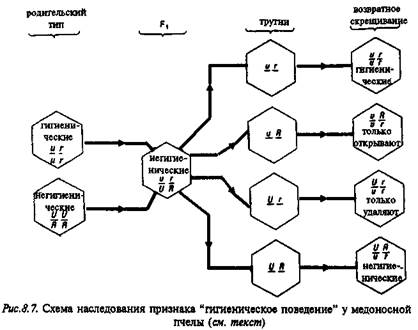
Генетический анализ – оценка поведения гибридов, полученных от скрещивания устойчивых к болезни и подверженных заболеванию пчел, показал, что это "гигиеническое поведение" детерминировано двумя независимыми генами, причем аллели, определяющие устойчивость к заболеванию, рецессивны (рис. 8.7).
Полученные в этом элегантном исследовании данные и сегодня остаются одним из немногочисленных примеров олигогенного наследования признаков поведения. Более подробно эти опыты изложены в учебнике Д.Мак-Фарленда (1988).
Движения пчел, которые они совершают при вскрытии ячеек и выбрасывании зараженных личинок, относятся к категории ФКД и обладают всеми характерными для них признаками. Данные движения выполняются насекомыми в полной форме без предварительного опыта и практически неизвестно, чтобы они видоизменялись при "тренировке". Более того, эти движения неспецифичны и характерны для вида в целом, а не только для каких-либо пород пчел. Все пчелы выполняют эти движения, однако у пород, подверженных заболеванию, частота этих ФКД чрезвычайно низка. Поэтому их поведение не может играть своей адаптивной роли: зараженные личинки остаются, и инфекция распространяется.
 Генетические исследования фуражировочного поведения медоносной пчелы были проведены на селектированных линиях. Селекцию проводили по признаку "большое или малое количество собираемой пыльцы". После получения пятого поколения селекции провели скрещивание двух линий и проанализировали поведение гибридов F1 и F2, а также беккроссов.
Генетические исследования фуражировочного поведения медоносной пчелы были проведены на селектированных линиях. Селекцию проводили по признаку "большое или малое количество собираемой пыльцы". После получения пятого поколения селекции провели скрещивание двух линий и проанализировали поведение гибридов F1 и F2, а также беккроссов.
Помимо классического генетического анализа был также использован метод картирования "локусов количественных признаков" (см.: 8.6.6.4), позволяющий с помощью специальных молекулярных маркеров выявлять гены, участвующие в определении данного признака, т.е. находить их положение на той или иной хромосоме.
Оказалось, что количество пищи, собираемое пчелой, детерминировано двумя локусами – pln-1 и рЫ-2 – в группах сцепления Пи X.
Генетические данные, полученные как характеристика особенностей фуражировочного поведения колоний, были подтверждены при исследовании поведения отдельных особей. Особи-фуражиры из линии с высокими показателями сбора пищи всегда приносили больше нектара или пыльцы, чем особи из линии с низкими значениями признака.
ПАДАЛЬНАЯ МУХА
Генетика падальной мухи (Phormia regina) изучена очень слабо, в особенности по сравнению с дрозофилой. В то же время крупные размеры этого насекомого позволили Дж. Хиршу (J. Hirsch) и его коллегам еще в 70-е годы провести на этом объекте исследования по генетике обучения пищевой реакции, которые и по сей день остаются уникальными (Me Guire, Hirsoh, 1977).
Спомощыо специальной пипетки насекомое фиксируется так, чтобы можно было наблюдать, как оно вытягивает хоботок (рис. 8.8).
 Безусловной реакцией, на основе которой вырабатывается условный рефлекс, является вытягивание хоботка при нанесении на оральную область 0,5М раствора сахарозы. Нижние членики передней лапки, где расположены хемочувствительные нервные окончания, помещают либо в дистиллированную воду, либо в раствор какого-либо вещества (но не сахара). При такой схеме эксперимента у насекомого вырабатывается условная реакция вытягивания хоботка в ответ на хемосигналы, ранее не вызывавшие такой реакции. Однако, как это всегда бывает при развитии процесса обучения у примитивных животных, одновременно выявляется сильное влияние процесса сенси-тизации, т.е. изменения чувствительности эффекторной системы не за счет формирования ассоциативной связи, а как результат совместного действия ряда стимулов и возбуждения насекомого в результате фиксации. При исследовании пищевых реакций Phormia regina процесс сенситизации исследователи назвали центральным возбужденным состоянием (central exitatory state – CES).
Безусловной реакцией, на основе которой вырабатывается условный рефлекс, является вытягивание хоботка при нанесении на оральную область 0,5М раствора сахарозы. Нижние членики передней лапки, где расположены хемочувствительные нервные окончания, помещают либо в дистиллированную воду, либо в раствор какого-либо вещества (но не сахара). При такой схеме эксперимента у насекомого вырабатывается условная реакция вытягивания хоботка в ответ на хемосигналы, ранее не вызывавшие такой реакции. Однако, как это всегда бывает при развитии процесса обучения у примитивных животных, одновременно выявляется сильное влияние процесса сенси-тизации, т.е. изменения чувствительности эффекторной системы не за счет формирования ассоциативной связи, а как результат совместного действия ряда стимулов и возбуждения насекомого в результате фиксации. При исследовании пищевых реакций Phormia regina процесс сенситизации исследователи назвали центральным возбужденным состоянием (central exitatory state – CES).
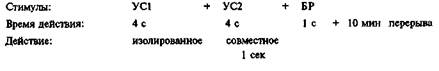
Чтобы отделить феномен формирования истинной ассоциативной связи от явлений CES, была разработана специальная схема эксперимента.
При такой последовательности стимулов двигательная безусловная реакция (вытягивание хоботка) в ответ на УС1 является отражением CES, т.е. следствием увеличения чувствительности к безусловному стимулу. Реакция же на УС2 отражает формирование условной связи; 10-минутный перерыв после сочетания стимулов способствовал угасанию CES. К. 15-му применению комплекса стимулов доля мух с условной реакцией росла, что свидетельствовало о формировании ассоциативной связи.
С помощью искусственного отбора на быстрое и медленное образование ассоциативной связи получили две линии мух, различия между которыми по исследуемому признаку постепенно увеличивались в процессе селекции. Искусственный отбор на быстроту формирования CES дал иную картину динамики различий:уже после первого поколения отбора селектируемые линии стали резко различаться по быстроте формирования CES, однако дальше по ходу селекции эти различия оставались неизменными (рис. 8.9).
Столь разные ответы на отбор у двух признаков свидетельствуют о разном характере их генетической детерминации. Быстрый ответ с последующим параллельным ходом кривых (как в случае CES) свидетельствует о моногенном наследовании признака, тогда как постепенное расхождение кривых, соответствующих уровням признака в разных поколениях, – следствие его полигенной детерминации.
Поскольку падальная муха – достаточно крупный объект, который позволяет вести нейрофизиологические исследования с меньшими трудностями, чем, например, на дрозофиле, результаты селекционных экспериментов представляются особенно интересными для изучения нейробиологии классического условного рефлекса.
Исследование генетических закономерностей в проявлении физиологических признаков у "простых" объектов, информация о дифференциальной экспрессии нейрогенов в корреляции с разными функциями существенно помогают в формировании новых подходов к анализу генетики поведения у более сложных, но традиционных объектов – дрозофилы и лабораторных грызунов.
8.4.3. ДРОЗОФИЛА
8.4.3.1.ПОВЕДЕНЧЕСКИЙ репертуар дрозофилы. Характерной чертой поведения насекомых (в том числе мух рода Drosophila) является преобладание в нем видоспецифических фиксированных комплексов действий. Однако наряду с ними в поведенческом репертуаре плодовой мушки имеются как более простые (таксисы), так и более сложные (реакции, приобретенные при обучении) феномены. Наиболее подробно изучено поведение Drosophila melanogaster.
Биологические ритмы. Характерный для дрозофилы ритм суточной активности обнаруживается не столько в уровне локомоции, сколько в частоте вылупления взрослых особей из куколок. Такие изменения в физиологических функциях очевидным образом модулируются чередованием света и темноты, однако, как это показано и для других животных, существуют также и внутренние механизмы поддержания околосуточных колебаний.
Фото-, гео- и другие таксисы. Это наиболее четкие, относительно примитивные реакции, выражающиеся в виде упорного перемещения насекомых в градиенте интенсивности света, концентрации химического вещества, геомагнитного поля и др. На стремлении мух перемещаться к свету (фототаксис) базируются методики исследования способности дрозофилы обучаться избегать неприятного запаха при его сочетании с ударом электрического тока.
Пищевое поведение. В ответ на вкусовое раздражение у мухи вытягивается хоботок. Эта безусловная реакция легла в основу выработки пищевого условного рефлекса у падальной мухи. Проведение таких исследований у плодовой мушки затруднено в связи с ее малыми размерами, однако известно, например, что эта реакция сильно различается у самцов и самок. Более плодотворны проводимые на дрозофиле исследования генетического контроля пищевого предпочтения. Эту особенность поведения изучают как на личинках (поиск пищи), так и на взрослых особях в связи с выбором самкой пищевого объекта для откладки яиц.
Реакция на обонятельные раздражители. Особый аспект пищевого поведения мух представляет собой их положительная реакция на обонятельные сигналы – пищевые аттрактанты. Подобные реакции осуществляются по механизмам таксисов, однако конкретных данных об их физиологических механизмах немного. Эти таксисы достаточно часто используются в экспериментах по генетике поведения, для которых такая информация необходима. В частности, положительная реакция на пищевой аттрактант не обязательно бывает связана с высоким уровнем локомоторной активности.
Оборонительное поведение. Обонятельные сигналы могут вызывать и реакции избегания. К их числу относится бензальдегид, широко используемый в экспериментах с обучением дрозофилы. В качестве отрицательного воздействия, на основе которого можно сформировать условную реакцию избегания, используется раздражение электрическим током. Физиологические корреляты такого воздействия изучены мало.
Половое поведение. У двукрылых, и в частности у дрозофилы, этот вид поведения организован достаточно сложно. Собственно спариванию предшествует более или менее интенсивно выраженное поведение ухаживания, функция которого – активация обоих партнеров (см.: рис. 8.1). Процесс ухаживания включает в себя несколько стадий. Первая из них – это стадия ориентации, когда самец занимает перед самкой положение, перпендикулярное продольной оси ее тела. Во время второй стадии он осуществляет вибрацию крылом, повернутым в горизонтальной плоскости на 90°. Эта фаза очень вариабельна по длительности (она укорочена при мутации yellow) и по "рисунку" отдельных пачек сигналов, а также интервалов между ними. Затем следуют стадии облизывания гениталии и собственно садки.
После копуляции у самки развивается временная половая невосприимчивость, а в ее организме образуется химическое вещество, вызывающее у самцов реакцию избегания. Это специфическое вещество цкс-вакцинилацетат называется антиафродитным феромоном. Генетические исследования полового поведения некоторых мутантов (см.: 8.4.3.5) показали, что самец, который сделал неудачную попытку ухаживания за невосприимчивой самкой, в дальнейшем, в течение примерно 2 ч воздерживается от ухаживания даже за рецептивными самками. Иными словами, мухи обнаруживают способность к торможению поведенческой реакции вследствие неприятного раздражения, т.е. они обучаются после единственного сочетания условного (вид самки) и безусловного (антиафродитный феромон) сигналов.
Обучение. Способность дрозофилы к обучению реакции избегания электрического тока исследуется в специальных установках с помощью методики "обратного потока" (см.: 8.4.3.6). Этот метод позволяет быстро тестировать большое число особей, что особенно важно при поиске мутантов с измененным поведением.
Очевидное адаптивное значение в жизни дрозофилы в естественных условиях имеет недавно описанная способность этих насекомых обучаться подавлению локомоторной активности. В экспериментах, проведенных в Институте физиологии им. Павлова РАН Н.Г. Камышевым с соавт. (1994), четко продемонстрировано, что исходный уровень локомоторной активности мухи достоверно снижается, когда ее подсаживают к группе из 10 других мух, причем это снижение нельзя объяснить ни реакцией на какие-либо вещества, ни безусловнорефлекторным торможением активности, ни привыканием.
Снижение двигательной активности мух выражается, в частности, в укорочении длительности отдельных "пробежек" и снижении числа контактов особей между собой. Оно имеет условнорефлек-торную природу и является результатом формирования инструментального навыка подавления активности в результате отрицательного опыта: при контактах мухи обмениваются "ударами" конечностей, что играет роль наказания. В естественных условиях, при скоплении дрозофил на источнике пищи – гниющих фруктах – способность к подобному обучению, видимо, обеспечивает оптимальное использование пищевых ресурсов, а это, несомненно, способствует выживанию.
Не нашли, что искали? Воспользуйтесь поиском: