ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
ФИЗИКО-ХИМИЧЕСКИЕ ПРОЦЕССЫ, ПРОТЕКАЮЩИЕ ПРИ МЕТАБОЛИЗМЕ МИКРООРГАНИЗМОВ В ХЛЕБОПЕКАРНЫХ ПОЛУФАБРИКАТАХ
В технологии хлебобулочных изделий для обеспечения процессов спиртового брожения и гликолиза применяют хлебопекарные прессованные дрожжи и молочнокислые бактерии, содержащиеся в бактериальных хлебопекарных заквасках. Микробиологическим процессам, протекающим под их действием в полуфабрикатах, присущи огромные потенциальные возможности.
Отличительной особенностью дрожжевой клетки является то, что структуры высшего порядка белков и нуклеиновых кислот в ней определяют ее метаболизм. В состав дрожжевой клетки входят биологически важные макромолекулы, сложная организация и воспроизведение которых способствуют ее жизнедеятельности. Следует отметить и тот факт, что все компоненты клетки (белки, нуклеиновые кислоты, липиды, углеводы) непрерывно изменяются, постоянно обновляются.
Метаболизм дрожжевых клеток и бактерий зависит от их способности пропускать воду и растворенные в ней вещества, т.е. зависит от их проницаемости. Эта способность неодинакова у дрожжевых клеток и молочнокислых бактерий. Проблема проницаемости имеет большое значение в механизме обмена веществ, в вопросах распределения веществ между клетками, возникновения биоэлектрических потенциалов.
В основе проникновения вещества в клетку лежит процесс диффузии, математическую трактовку которого дал Фик (1955). По закону Фика, скорость диффузии dm/dt прямо пропорциональна градиенту концентрации dc/dx и площади поверхности s, через которую осуществляется диффузия:

|
Коэффициент пропорциональности D в уравнении носит название константы диффузии.
Скорость проникновения вещества в клетку определяется по формуле, предложенной Коллендером и Берлундом:
 , где
, где
С-С0–разность концентраций вещества между клеткой и средой
K– константа проницаемости
Вычисление константы проницаемости проводят по формуле,которую можно представить в следующем виде:
Принимая 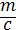 = v, (v — объем клетки), и интегрируя, получаем:
= v, (v — объем клетки), и интегрируя, получаем:

Считая v и s постоянными в течение всего времени эксперимента, получим:

|
Проникновение вещества может происходить и в направлении, обратном концентрационному градиенту, при наличии другого градиента, противоположно направленного, например, электрического.
При исследовании проницаемости следует постоянно учитывать роль метаболических процессов и в обеспечении концентрационного градиента между клеткой и средой. Так, включение поступающего в клетку аниона фосфора в молекулы нуклеотидов, нуклеиновых кислот, нуклеопротеидов и фосфолипидов приводит к снижению концентрации неорганического фосфора в клетке и тем самым поддерживает градиент этого аниона между клеткой и средой.
Важную роль в накоплении веществ в клетке играет и химическая адсорбция.
Направление потока веществ не остается раз навсегда постоянным: оно может измениться на обратное при нарушении соотношений между процессами синтеза и распада важнейших высокомолекулярных компонентов протоплазмы. Так, в молодых клетках происходит накопление ионов калия и фосфатов за счет включения последних в молекулы АТФ. Напротив, по мере старения клеток происходит распад органических фосфатов с отщеплением и последующим выходом фосфатов, а также ионов калия в среду. Иначе говоря, направление потока веществ изменяется на противоположное.
При всем разнообразии строения и физико-химических свойств молекул веществ существуют в основном два пути их проникновения в клетку:
1) через субмикроскопические заполненные водой поры, пронизывающие протоплазму и соединяющие ее с внешней средой, проникают молекулы водорастворимых веществ и ионы;
2) за счет растворения в липидах, входящих в состав протоплазмы и особенно ее поверхностных слоев, проникают водонерастворимые органические соединения.
Проницаемость клеток для органических молекул уменьшается по мере возрастания количества гидроксильных, карбоксильных и аминогрупп.
Для диффузии молекул водорастворимых соединений, при условии если их радиус не превышает радиуса гексоз, применимо уравнение Товерта:
 , где
, где
D – константа диффузии
М – молекулярная масса диффундирующего вещества
Джекобс (М. Jacobs), исходя из современной теории электрического строения молекул, показал, что все химические соединения, а также отдельные, входящие в их состав радикалы, можно разбить на две большие группы:
1) электрически симметричные, т.е., у которых центры положительных и отрицательных зарядов совпадают друг с другом, названные неполярными, или гомеополярными (метиловые, этиловые, фениловые группы);
2) электрически несимметричные, центры положительных и отрицательных зарядов которых не совпадают, - полярные или гетерополярные (соединения с группами СООН, ОН и NH2).
Неполярные соединения хорошо растворимы в липидах, хлороформе и ацетоне, имеют низкую диэлектрическую постоянную, а полярные хорошо растворимы в воде и характеризуются высокой диэлектрической постоянной.
Исключение из этого правила составляет глюкоза, плохо растворимая в липидах, но легко проникающая в клетку. Возможно, что глюкоза нарушает глобулярную структуру некоторых белков. Аномально высокая проницаемость клеток для глицерина может быть отнесена на счет его способности образовывать димеры, в которых маскирующие друг друга полярные гидроксильные группы соседних молекул не препятствуют липидорастворимости.
Поступление веществ из внешней среды обеспечивают клеточная стенка дрожжевой клетки, цитоплазматическая мембрана, пиноцитоз и эндоплазматическая сеть.
Бактерии имеют относительно простое строение. Бактериальная клетка состоит из протоплазмы (цитоплазмы) с различными включениями, которая от внешней среды отделена плотной оболочкой — клеточной стенкой. Стенки бактериальных клеток представляют собой жесткие, пористые, замкнутые структуры, обеспечивающие клеткам защиту от физических воздействий, препятствующие набуханию и разрыву клеточной мембраны. Клеточные стенки бактерий принимают участие во многих физиологических и физико-химических процессах, выполняя различные функции в жизни клетки. Оболочки клеток химически активны, в них происходит множество химических реакций. Цитоплазматическая мембрана бактерий контролирует поступление питательных веществ и выход продуктов метаболизма из клеток.
Физико-химические процессы проницаемости дрожжевых клеток для неполярных соединений объясняются липидной структурой клеточной мембраны. Липид ведет себя как растворитель для неполярных участков белков, стероидов и т.д. В цитоплазматической мембране белковые молекулы, расположенные между слоями липидов, могут изменять свое положение. Липиды, согласно глобулярной структуре биомембраны дрожжей, организованы в биослои, но они не являются непрерывными (рис. 6.4).

Толщину биомолекулярного слоя можно вычислить по формуле Фрикке:
 r
r  , где
, где
С- ёмкость клеток, найденная в опыте
S – суммарная площадь поверхности клеток
 – диэлектрическая проницаемость оболочки, равная для липидов 2-3
– диэлектрическая проницаемость оболочки, равная для липидов 2-3
r – пересчётный коэффициент, равный 8,84 * 10-14 , для случаев, когда емкость дана в Фарадах, а размеры в см
d – искомая толщина оболочки (cм)
Расчеты, произведенные на основании опытных данных, показали, что толщина клеточной мембраны приблизительно равна 32 А, что соответствует бимолекулярному слою жирных кислот с длиной углеводородной цепочки в 16 - 18 углеродных атомов.
В клетку могут проникать вещества, нерастворимые в липидах, в первую очередь, минеральные ионы, сахара и аминокислоты.
Для того чтобы молекула или ион водорастворимого вещества проникли в клетку, они должны обладать некоторым добавочным количеством энергии. Пересекая клеточную мембрану, они должны, во-первых, оторваться от окружающей их сольватной оболочки, во-вторых, раздвинуть плотно расположенные молекулы поверхностного слоя на границе клетка — омывающий ее раствор, в-третьих, преодолеть взаимодействие своих полярных групп с полярными группами, расположенными внутри канала поры, в-четвертых, преодолеть энергетический барьер на границе поверхностной мембраны и водного раствора коллоидов и ионно-дисперсных веществ протоплазмы.
Изменяя активную реакцию внешней среды, можно увеличить или уменьшить количество недиссоциированных молекул и тем усилить или ослабить проникновение вещества в клетку.
Процессы обмена веществ в дрожжевой клетке протекают с участием биологических катализаторов, в состав которых входят экзо- и эндоферменты. Взаимосвязь дрожжевых клеток с окружающей средой, приспособление к ней создают благоприятные физико-химические условия и предопределяют активность их ферментов, ускоряют и обеспечивают внутриклеточные биохимические превращения.
В условиях хлебопекарного производства факторами, влияющими на рациональный ход технологического процесса и качество продукции, являются исходная биологическая активность дрожжевых клеток и способность их усваивать питательные вещества. Эффективность их усвоения зависит от ряда физико-химических факторов: проницаемости клеточной мембраны, возникновения биоэлектрических потенциалов, величин концентрационного и электрического градиентов, наличия водо- и нерастворимых веществ в окружающей среде, рН среды, величины энергетического барьера, активности и др. Уникальным свойством дрожжевой клетки является непрерывное получение энергии - химической, электрической, необходимой для ее метаболизма и физиологических функций. Энергия в клетке накапливается в результате окислительно-восстановительных реакций.
Используя научные концепции метаболизма дрожжей, коэффициент распределения питательных веществ между клетками и окружающей средой, можно регулировать их активность при брожении хлебопекарных полуфабрикатов.
Не нашли, что искали? Воспользуйтесь поиском: