ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Потенціал дії. Механізм виникнення та поширення нервового імпульсу
Розглянуті потенціали спокою свідчать про те, що цитоплазма незбудженої клітини має негативний електричний потенціал по відношенню до позаклітинного середовища  Збудження клітини може спричинити виникнення потенціалу дії (або спайку), при цьому спостерігається спонтанне обернення знака мембранного потенціалу.
Збудження клітини може спричинити виникнення потенціалу дії (або спайку), при цьому спостерігається спонтанне обернення знака мембранного потенціалу.
Для виникнення потенціалу дії (ПД), необхідно викликати локальний зсув потенціалу  на внутрішній поверхні мембрани до значення, яке перевищувало б значення, що називається пороговим (тобто
на внутрішній поверхні мембрани до значення, яке перевищувало б значення, що називається пороговим (тобто  ). Таку зміну потенціалу
). Таку зміну потенціалу  можна викликати штучно, пропускаючи крізь мембрану короткочасний збуджуючий струм (стимул). Якщо зсув негативного потенціалу на внутрішньому боці мембрани не перевищує
можна викликати штучно, пропускаючи крізь мембрану короткочасний збуджуючий струм (стимул). Якщо зсув негативного потенціалу на внутрішньому боці мембрани не перевищує  то після вимкнення стимулу потенціал
то після вимкнення стимулу потенціал  повільно повертається до первісного значення (рис. 4.33а,б).
повільно повертається до первісного значення (рис. 4.33а,б).
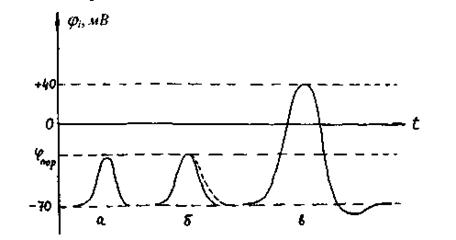
Рис. 4.33. Збудження мембрани за допомогою короткочасного збуджуючого струму (стимулу): а - підпороговий імпульс; б - пороговий імпульс (згасаючий спайк); в - потенціал дії (ПД).
Якщо ж зсув  перевищив
перевищив  , то виникне подальше спонтанне зростання
, то виникне подальше спонтанне зростання  і після припинення стимулу. При цьому значення
і після припинення стимулу. При цьому значення  досягає нуля і продовжує швидко мінятися у додатному напрямку (рис. 6.ЗЗв). Таким чином, виникає обернення знака
досягає нуля і продовжує швидко мінятися у додатному напрямку (рис. 6.ЗЗв). Таким чином, виникає обернення знака  (деполяризація мембрани). Потенціал
(деполяризація мембрани). Потенціал  досягає максимального значення
досягає максимального значення  після чого протягом приблизно мілісекунди, повертається до вихідного значення
після чого протягом приблизно мілісекунди, повертається до вихідного значення  В аксоні кальмара весь спайк триває близько 6 мс.
В аксоні кальмара весь спайк триває близько 6 мс.
Виникнення потенціалу дії спричинене різким збільшенням проникності мембрани для іонів  , що виникає за умови зростання потенціалу
, що виникає за умови зростання потенціалу  вище
вище  . Іони
. Іони  входячи всередину клітини, призводять до ще більшого зростання потенціалу
входячи всередину клітини, призводять до ще більшого зростання потенціалу  на внутрішньому боці мембрани, що викликає, в свою чергу, подальше зростання проникності мембрани для іонів
на внутрішньому боці мембрани, що викликає, в свою чергу, подальше зростання проникності мембрани для іонів  і, як наслідок, - зростання швидкості їх проникнення всередину клітини Зростання потенціалу на внутрішньому боці мембрани викличе, з деяким запізненням, повільне збільшення проникності мембрани для іонів
і, як наслідок, - зростання швидкості їх проникнення всередину клітини Зростання потенціалу на внутрішньому боці мембрани викличе, з деяким запізненням, повільне збільшення проникності мембрани для іонів  внаслідок пасивної дифузії вони почнуть виходити назовні.
внаслідок пасивної дифузії вони почнуть виходити назовні.

Рис. 4.34. Ілюстрація механізму формування потенціалу дії: 1 - деполяризація; 2 - реполяризація.
При цьому спостерігається повернення мембранного потенціалу  від максимального до вихідного значення (від +40 до -70 мВ). Це процес так званої реполярізації (див. рис. 4.34).
від максимального до вихідного значення (від +40 до -70 мВ). Це процес так званої реполярізації (див. рис. 4.34).
Іони  , що увійшли у клітину під час зростання ПД, можуть вийти зовні тільки внаслідок роботи
, що увійшли у клітину під час зростання ПД, можуть вийти зовні тільки внаслідок роботи  -насоса.
-насоса.
Після ПД спостерігається період незбудження або абсолютний рефракторний період, під час якого не може виникнути наступний ПД. Він триває від 0.5 до 2 мс. За цей час внаслідок роботи  - насоса відбувається відновлення вихідних значень концентрацій іонів
- насоса відбувається відновлення вихідних значень концентрацій іонів 
Таким чином, формування ПД спричинене двома іонними потоками крізь мембрану: потік іонів  всередину клітини викликає перезарядку мембрани, а протилежно спрямований потік
всередину клітини викликає перезарядку мембрани, а протилежно спрямований потік  - відновлення вихідного значення потенціалу спокою. Потоки зсунуті у часі, завдяки чому можлива поява ПД (рис. 4.35).
- відновлення вихідного значення потенціалу спокою. Потоки зсунуті у часі, завдяки чому можлива поява ПД (рис. 4.35).
Потенціал дії, що виник на певній ділянці нервової клітини, швидко розповсюджується вздовж її поверхні завдяки локальним струмам між збудженими і незбудженими ділянками нервового волокна (рис. 4.36). Локальні струми чинять подразнюючу дію на сусідні незбуджені ділянки і викликають зміну проникності мембрани. Локальні струми деполяризують мембрану до певного рівня, а ПД на кожній ділянці виникає внаслідок іонних потоків  спрямованих перпендикулярно напрямку поширення збудження.
спрямованих перпендикулярно напрямку поширення збудження.

Рис. 6.35. Вплив зміни проник- Рис. 6.36. Ілюстрація теорії ло-
ностей натрійових та калійових кальних струмів: 1 - немієлінізо-
каналів на формування потенці- ване волокно; 2 - мієлінізоване. алу дії.
Теорію генерації і поширення потенціалу дії запропонували у 1948-1952 рр. А. Ходжкін і А. Хакслі. Згідно з моделлю Ходжкіна і Хакслі зміна іонних провідностей при зсувах мембранних потенціалів спричинена впливом електричного поля на просторовий розподіл у мембрані заряджених активуючих частинок.
На рис. 4.37 подано еквівалентну схему елемента збудливої мембрани нервового волокна. Вона становить електричне коло з чотирма паралельними вітками. Одна з них містить електричну ємність, а інші відтворюють натрієву  калієву
калієву  провідності мембрани і так звану провідність відтоку
провідності мембрани і так звану провідність відтоку 

Рис.4.37. Еквівалентна електрична схема ділянки мембрани
У кожну з цих віток включені джерела електрорушійної сили  які дорівнюють відповідним рівноважним потенціалам Нернста:
які дорівнюють відповідним рівноважним потенціалам Нернста:

Вираз для електричного струму крізь мембрану можна подати у такому вигляді:
 (4.65)
(4.65)
де  Перший доданок у рівнянні (4.65) – це ємнісний струм, пов'язаний зі зміною різниці потенціалів на мембрані, а другий доданок - іонний струм.
Перший доданок у рівнянні (4.65) – це ємнісний струм, пов'язаний зі зміною різниці потенціалів на мембрані, а другий доданок - іонний струм.
Експериментально Ходжкін і Хакслі довели, що іонні струми  прямо пропорційні різниці потенціалів на мембрані
прямо пропорційні різниці потенціалів на мембрані  і рівноважному мембранному потенціалу Нернста для відповідного сорту іонів:
і рівноважному мембранному потенціалу Нернста для відповідного сорту іонів:
 (4.66)
(4.66)
Останнє рівняння в (4.66) враховує іонні струми всіх інших іонів, а саме:  тощо. Коефіцієнти
тощо. Коефіцієнти  в (4.66) характеризують провідності каналів. Провідність каналів може змінюватись внаслідок їх активації. Активація каналу - процес імовірнісний, котрий можна описувати за допомогою рівнянь хімічної кінетики за схемою:
в (4.66) характеризують провідності каналів. Провідність каналів може змінюватись внаслідок їх активації. Активація каналу - процес імовірнісний, котрий можна описувати за допомогою рівнянь хімічної кінетики за схемою:

де  - константи швидкостей активації та інактивації відповідно. Нехай
- константи швидкостей активації та інактивації відповідно. Нехай  - загальна кількість каналів, а
- загальна кількість каналів, а  -кількість активованих каналів, тоді
-кількість активованих каналів, тоді  - кількість неактивованих каналів. Швидкість виникнення активованих каналів буде такою:
- кількість неактивованих каналів. Швидкість виникнення активованих каналів буде такою:
 (4.67)
(4.67)
Поділимо рівняння (4.67) на  і позначимо через
і позначимо через  частку активованих каналів. Тоді рівняння (4.67) набуде вигляду
частку активованих каналів. Тоді рівняння (4.67) набуде вигляду
 (4.68)
(4.68)
або
 (4.69)
(4.69)
Розв'язок рівняння (4.69) має такий вигляд:
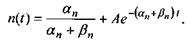
Константу  визначимо з такої початкової умови:
визначимо з такої початкової умови:  якщо
якщо  що дає
що дає

Остаточно маємо
 (4.70)
(4.70)
Коли  У результаті
У результаті
 (4.71)
(4.71)
де  - постійна часу встановлення рівноваги в каналі. Графік рівняння (4.71), що описує кінетику активації каналів, наведено на рис. 4.38. Експериментально було встановлено, що для калієвого каналу
- постійна часу встановлення рівноваги в каналі. Графік рівняння (4.71), що описує кінетику активації каналів, наведено на рис. 4.38. Експериментально було встановлено, що для калієвого каналу
 (4.72)
(4.72)
де  - максимальне значення провідності.
- максимальне значення провідності.
Ступінь "4" пов'язаний з тим, що  -канал відчиняється, коли до нього підходять чотири негативні заряди (наприклад, коли канал залишають два іони
-канал відчиняється, коли до нього підходять чотири негативні заряди (наприклад, коли канал залишають два іони  Для натрієвого каналу
Для натрієвого каналу
 (4.73)
(4.73)
де  - частка активованих, а
- частка активованих, а  - інактивованих каналів. Згідно з (4.73)
- інактивованих каналів. Згідно з (4.73)  -канал відчиняється, коли до нього потрапляють три активуючі (негативні) і видаляється одна блокуюча (позитивна) частинки.
-канал відчиняється, коли до нього потрапляють три активуючі (негативні) і видаляється одна блокуюча (позитивна) частинки.

Рис. 4.38. Кінетика активації каналів. Рис.4.39. Електричні струми
крізь елемент аксону.
Рівняння, які описують зміну в часу активованих та інактивованих  -каналів, мають той самий вигляд (4.68), що й для
-каналів, мають той самий вигляд (4.68), що й для  -каналів, а саме:
-каналів, а саме:
 (4.74)
(4.74)
Таким чином, рівняння для сили струму крізь мембрану (4.65) з урахуванням (4.66), (4.72) та (4.73) набуває такого вигляду:
 (4.75)
(4.75)
Знайдемо зв'язок між струмом крізь мембрану і струмом вздовж аксона. Для цього розглянемо елемент поверхні аксона довжиною  (рис. 4.39). Площа поверхні цього елемента
(рис. 4.39). Площа поверхні цього елемента  де
де  - радіус аксона. Сила
- радіус аксона. Сила
струму крізь мембрану аксона
 (4.76)
(4.76)
Збільшення мембранного струму  буде призводити до зменшення струму
буде призводити до зменшення струму  вздовж аксона, тобто
вздовж аксона, тобто
 (4.77)
(4.77)
З рівнянь (4.76) та (4.77) випливає, що
 (4.78)
(4.78)
Отже, сила струму в аксоплазмі згідно з законом Ома становить

де  - опір аксоплазми, причому
- опір аксоплазми, причому

де  - питомий опір аксоплазми. Тоді рівняння (4.78) набуває такого вигляду:
- питомий опір аксоплазми. Тоді рівняння (4.78) набуває такого вигляду:
 (4.79)
(4.79)
Остаточно, враховуючи (4.79), отримаємо рівняння Ходжкіна -Хакслї.
 (4.80)
(4.80)
У ліву частину рівняння Ходжкіна-Хакслі введемо швидкість поширення нервового імпульсу, беручи до уваги, що поширення нервового імпульсу вздовж аксона можна описати хвильовим рівнянням:

Тоді рівняння Ходжкіна-Хакслі (4.80) набуває вигляду
 (4.81)
(4.81)
Розв'язок рівняння (4.81) з використанням одержаних експериментально значень  дає можливість
дає можливість  розрахувати швидкість поширення нервового імпульсу. Наприклад, в гігантському аксоні кальмара вона становить
розрахувати швидкість поширення нервового імпульсу. Наприклад, в гігантському аксоні кальмара вона становить 
Як відомо, швидкість поширення ПД залежить від того, чи має аксон мієлінову оболонку, чи така оболонка відсутня. У першому випадку розповсюдження нервового імпульсу відбувається стрибками через перехвати Ранв'є і тому має значно більшу швидкість, ніж у другому випадку.
Не нашли, что искали? Воспользуйтесь поиском: