ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Гиперполяризационное торможение
Его суть заключается в том, что увеличивается потенциал мембраны по отношению к покою, в связи с чем снижается способность клетки реагировать на раздражители. Возникшее явление характеризуется понижением возбудимости (рис. 4.3). Одним из проявлений этого вида является положительный следовой потенциал, который возникает после потенциала действия и связан с тем, что еще часть ионов К+ не возвращена в клетку (на наружной поверхности мембраны остается достаточно большое количество ионов К+). В этом состоянии наблюдается явление относительной рефрактерности, т.е уменьшение ответа клетки на раздражитель. Необходим более сильный стимул, чтобы клетку возбудить. Из состояния гиперполяризации труднее достичь критического уровня деполяризации, чем из состояния покоя (рис. 4.3). Такое торможение чаще всего развивается в связи с большим выходом К+ наружу и большим поступлением Cl- в клетку. Изменение проницаемости мембран для К+ и Cl-, которое приводит к гиперполяризации мембраны, происходит под влиянием гамма - аминомасляной кислоты (ГАМК), вещества которое вырабатывают специальные нервные клетки (клетки Реншоу); серотонина и некоторых других веществ. Эти вещества увеличивают проницаемость мембраны для К+ и Cl- и приводят к гиперполяризации мембраны (более -100 мв) - формируется тормозной потенциал. Клетка переходит в состояние пониженной возбудимости.
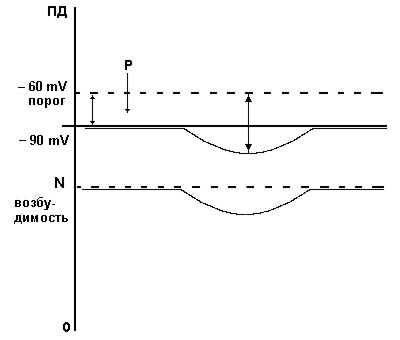
Рис. 4.3. Схема возникновения гиперполяризационного иорможения.
Работа нервной системы чаще всего связана с этим видом торможения. В зависимости от места возникновения торможения различают: пресинаптическое (А) и постсинаптическое (Б); центральное и периферическое (рис. 4.4).
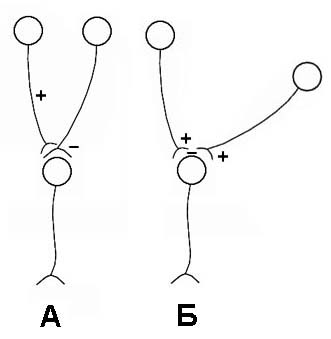
Рис. 4.4. Пресинаптическое (А) и постсинаптическое (Б) торможение
При пресинаптическом торможении чаще всего возникает деполяризация пресинаптической мембраны, в результате чего уменьшается выделение медиатора и передача импульса в синапсе. Постсинаптическое торможение развивается обычно по механизму гиперполяризационного и связано с возникновением тормозного постсинаптического потенциала (ТПСП) под влиянием нейромедиаторов (ГАМК, серотонина).
Центральное торможение имеет место в ЦНС, и впервые было описано И.М. Сеченовым в 1840 г. Накладывая кристаллик NaCl на зрительные бугры (таламус) мозга лягушки, он обнаружил увеличение времени двигательной рефлекторной реакции на раздражитель. Это послужило основанием для заключения, что в ЦНС имеются специфические тормозные центры, возбуждение которых кристалликом соли вызывает торможение в центрах спинномозговых рефлексов (рис. 4.5). Позднее было доказано, что одни и те же центры могут находиться в состоянии возбуждения и в состоянии торможения; а специфическими тормозными центрами могут являться клетки Реншоу, выделяющие тормозной медиатор - ГАМК.
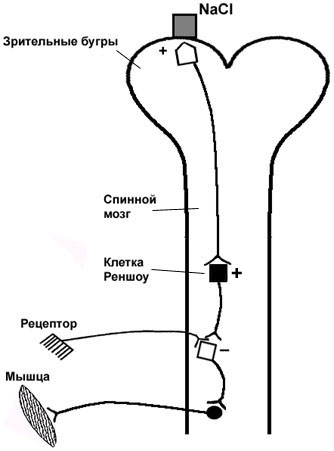
Рис. 4.5. Схема опыта "Сеченовское торможение"
Периферическое торможение было описано братьями Вебер на примере торможения деятельности сердца при раздражении блуждающего нерва. Периферическое торможение развивается вне ЦНС, в органах, и по механизму может быть как гиперполяризационное, так и деполяризационное.
Совокупность возбудительных и тормозных процессов в ЦНС и на периферии определяет специфическую биологически целесообразную деятельность нервной системы и различных органов.
Рецепторный потенциал
Возбуждение и торможение, которое возникает в нервной системе, создает все многообразие ответных реакций, обеспечивая процессы регуляции в организме. Существует специальная нервная структура, функция которой заключается в том, что она воспринимает различные раздражители и обеспечивает трансформацию любого вида энергии в нервный импульс. Эта структура называется рецептор. В рецепторе можно выделить две части: воспринимающую и трансформирующую.
Воспринимающая устроена таким образом, что стимул, действуя на нее, формирует в этих структурах генераторный (рецепторный потенциал). Это происходит вследствие растяжения мембраны клеток (проприорецепторы мышечной ткани), при действии механической волны (звуковые рецепторы), при изменении напряжения, давления (барорецепторы), при изменении электрохимического градиента (при действии химических веществ - хеморецепторы).
Если сила раздражителя и время действия увеличиваются, потенциал в этой части рецептора также увеличивается и удлиняется (рис. 4.6).
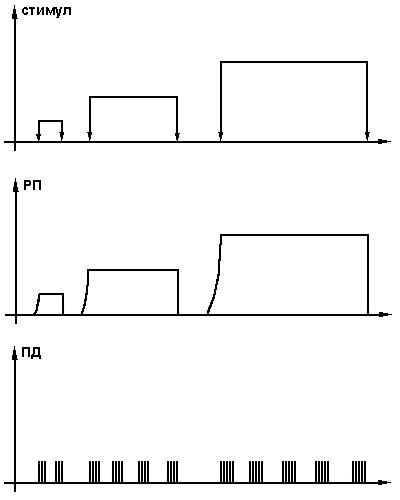
Рис. 4.6. Изменение рецепторного потенциала в зависимости от стимула.
При длительном действии раздражителя возникает явление привыкания (адаптации) рецептора, т.е уменьшается амплитуда потенциала.
Чем дольше действие раздражителя, тем быстрее наступает привыкание, чем сильнее раздражитель, тем хуже идет адаптация. Этот потенциал запускает во второй части рецептора формирование потенциала действия (ПД). Потенциал действия по уровню деполяризации везде одинаков. Он отличается по количеству импульсов, длительности интервалов между импульсами. Это явление называется трансформация рецепторного потенциала в потенциал действия. Рецепторный потенциал, как и местный, не распространяется (имеет локальный характер). Потенциал действия отвечает по принципу "все или ничего", обладает способностью к распространению. На выходе из рецепторов регистрируется ПД, который легко распространяется по нерву. В рецепторе создается частотный (сенсорный) код, который характеризует данный раздражитель. В зависимости от частоты и силы раздражителя, он будет отличаться количеством, частотой и длительностью импульсов, но не будет отличаться амплитудой (она одинакова). Любой по количеству стимул, действуя на соответствующий рецептор, трансформируется в ПД. Благодаря этому в НС по волокнам распространяются биопотенциалы с разной частотой и длительностью.
o 21. Роль нейромедиаторов и синаптических рецепторов в генерации возбуждающих и тормозных постсинаптических потенциалов.
Дубинин
o 3.4. Постсинаптические потенциалы. Распространение потенциала действия по нейрону
Запуск импульсной активности в нервной системе осуществляют два основных фактора. Первый из них — стимулы, действующие на чувствительные клетки сенсорных систем и изменяющие проницаемость их мембраны. Это приводит к развитию особых рецепторных потенциалов и в итоге — к генерации ПД.
Второй фактор — выделение медиатора из пресинаптического окончания. Попав в синаптическую щель, медиатор воздействует на постсинаптическую мембрану, возбуждая или тормозя следующий нейрон.
Процессы подобного возбуждения или торможения связаны с деятельностью еще одного типа ионных каналов — лиганд-зависимых (хемочувствительных). Они находятся на мембране, непосредственно окружающей синаптический контакт. Обычно они закрыты. Их открывание происходит лишь при появлении медиатора, несущего сигнал химического вещества (отсюда термин «хемочувствительные»).
Лиганд-зависимые каналы можно разделить на три основных класса: избирательно проницаемые по отношению к ионам Na+, ионам К+ и ионам Cl−. Отрывание первых из них приведет к входу в клетку ионов Na+ и деполяризации нейрона (рис. 3.14, а), во время которой разность потенциалов на мембране оказывается приближенной к порогу запуска ПД. В этот момент меньший, чем обычно, стимул может вызвать реакцию нейрона, т. е. нервная клетка находится в относительно возбужденном состоянии. В связи с этим локальная деполяризация мембраны под действием медиатора была названа возбуждающим постсинаптическим потенциалом (ВПСП). Медиаторы, вызывающие ВПСП, отнесены к группе возбуждающих медиаторов.
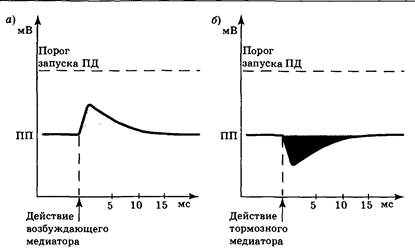
|
| Рис. 3.14.Изменение разности потенциалов на мембране нервной клетки при возникновении ВПСП (а) и ТПСП (б) |
Открывание хемочувствительных Cl−-каналов приводит к входу в клетку ионов хлора; открывание К+-каналов — к выходу ионов калия. В этих случаях возникает гиперполяризация и разность потенциалов на мембране нейрона увеличивается по абсолютной величине (рис. 3.14, б), поэтому для запуска ПД необходим больший, чем обычно, стимул. Следовательно, нервная клетка находится в относительно заторможенном состоянии. В связи с этим локальная гиперполяризация мембраны под действием медиатора была названа тормозным постсинаптическим потенциалом (ТПСП). Медиаторы, вызывающие ТПСП, отнесены к группе тормозных медиаторов.
Усредненные параметры ВПСП и ТПСП весьма близки (рис. 3.14). Их длительность составляет обычно около 10 мс (иногда 50—100 мс), что существенно больше, чем в случае ПД. Амплитуда ВПСП и ТПСП определяется длительностью и крутизной наклона их первой фазы, которая зависит от количества и длительности существования медиатора в синаптической щели. Амплитуда одиночных постсинаптических потенциалов в ЦНС составляет 1—5 мВ. В крупном нервно-мышечном синапсе аналог ВПСП — потенциал концевой пластинки, может достигать 40 и более мВ.
При детальном анализе сигнала можно видеть, что первые фазы ВПСП и ТПСП имеют ступенчатый характер, т. е. нарастают дискретно, шагами (квантами). Такая дискретность связана с тем, что выброс медиатора в синаптическую щель также идет квантами, где квант — одна везикула. В каждой везикуле содержится несколько тысяч молекул медиатора, и их воздействие на постсинаптическую мембрану вызывает сдвиг потенциала примерно на 0,1 мВ.
В подавляющем большинстве случаев (кроме потенциала концевой пластинки) одиночный ВПСП не способен запустить ПД, так как возбуждение, вызываемое медиатором, не дорастает до порогового уровня. Для достижения порога запуска ПД необходима суммация (наложение) нескольких ВПСП.
Выделяют два варианта суммации — временную и пространственную. Временная суммация — объединение эффектов стимулов, пришедших по одному «каналу» с большой частотой (рис. 3.15): если к еще не угасшему ВПСП присоединить второй, затем третий и т.д., — возникнет реальная возможность запустить ПД. Это означает, что сигнал, достигший синапса, достаточно интенсивен и «заслуживает» того, чтобы быть переданным дальше по сети нейронов.
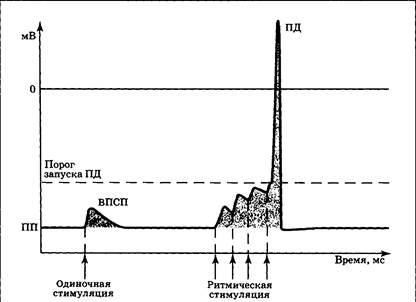
|
| Рис. 3.15. Схема временной суммации ВПСП, возникающих при повторной стимуляции синапса, выделяющего возбуждающий медиатор |
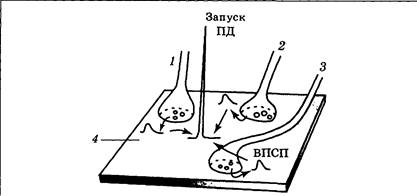
|
| Рис. 3.16. Схема пространственной суммации ВПСП в результате одновременного срабатывания синапсов 1 — 3 |
Пространственная суммация заключается в наложении друг на друга ВПСП соседних синапсов 1 — 3 в некоторой близлежащей точке постсинаптической мембраны 4 (рис. 3.16), обладающей потенциал-зависимыми ионными каналами. Схема пространственной суммации напоминает логическую ячейку по типу «И», т. е. результат положительный, если несколько условий будут выполнены (несколько входных сигналов одновременно достигнут нервной клетки).
В ходе деятельности нейронов эффекты пространственной и временной суммации объединяются, и чем больше синапсов участвуют в этом процессе (срабатывают относительно одномоментно), тем больше вероятность достичь порога запуска ПД. При этом часть синапсов может обладать тормозными свойствами и вызывать ТПСП, вычитающиеся из суммы возбуждающих влияний. В результате в первом приближении условие запуска ПД в каждый момент времени можно определить следующим образом:
ПП + (сумма всех ВПСП) − (сумма всех ТПСП) > порог запуска ПД
Не нашли, что искали? Воспользуйтесь поиском: