ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Передача наследственной информации в процессе размножения клеток
Задание 2.1 Изучить закономерности клеточного цикла, процессы протекающие в различные его периоды.
В основе всех видов размножения лежит деление клетки. Клетка претерпевает ряд последовательных изменений, из которых складывается клеточный цикл. Клеточный цикл состоит из четырех стадий (рис. 5):

Рисунок 5. Схема клеточного цикла:
G1 – пресинтетическая стадия; S – стадия синтеза; G2 – постсинтетическая стадия; М – деление (митоз или мейоз).
Пресинтетический (G1) период обычно самый продолжительный и следует за телофазой митоза. В этот период клетка интенсивно синтезирует РНК и белки, увеличивает свой объем, выполняет специфические функции, дифференцируется. На этой стадии хромосома представлена одной хроматидой
Если G1-период затягивается, то клетки могут прекратить делиться и перейти в так называемый G0-период. Клетки могут находиться в G0-периоде длительное время, начинают расти, дифференцироваться, достигая состояния терминальной (окончательной) дифференцировки. В этом случае окончание клеточного периода сопровождается гибелью клетки.
S-период (синтетический) – в клетке продолжается синтез белка, но главным в нем является то, что происходит удвоение (репликация) ДНК. Начинается удвоение центриолей в клеточном центре.
Постсинтетический (G2) период. Клетка готовится к делению, и идет остаточный синтез белка. Хромосома представлена двумя хроматидами, центриоли достигают размеров зрелых органелл. В этой фазе накапливается АТФ для энергетического обеспечения митоза.
Длительность клеточного цикла у разных организмов не одинакова.
Задание 2.2 Изучить механизмы репродукции и дифференцировки соматических клеток.
Митоз – деление, которое обеспечивает постоянство набора хромосом, то есть в его результате формируются две идентичные материнской и между собой по генотипу клетки.
Митотический цикл – это существование клетки от деления до следующего деления. Клеточный цикл может совпадать или не совпадать с митотическим. Если клеточный цикл совпадает с митотическим, то он состоит из митоза и интерфазы (рис. 6).
Интерфаза занимает основное время в жизнедеятельности клетки. В интерфазу клетка осуществляет функции межклеточной кооперации и поддержание жизни целого многоклеточного организма. В интерфазе последовательно различают фазы G1, S и G2.

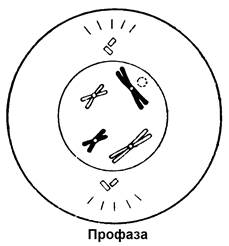
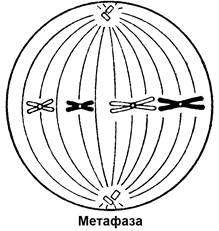


Рисунок 6. Схематическое изображение стадий митоза
Профаза. Самая продолжительная фаза клеточного деления. Хроматиды укорачиваются и утолщаются в результате их спирализации и конденсации. Хроматиды тесно ассоциированы по длине и перекручены. К концу профазы эта ассоциация ослабевает. Центриоли расходятся к противоположным полюсам клетки. От каждой центриоли в виде лучей расходятся короткие микротрубочки, образующие в совокупности звезду.
Ядрышки уменьшаются, так как их нуклеиновая кислота частично переходит в определенные пары хроматид. К концу профазы ядерная мембрана распадается и образуется веретено деления.
Прометафаза. Пары хроматид прикрепляются своими центромерами к нитям веретена (микротрубочкам) и перемещаются вверх и вниз по веретену до тех пор, пока их центромеры не выстроятся по экватору веретена перпендикулярно его оси. Прометафаза завершается, когда хромосомы достигают экваториальной плоскости.
Метафаза. В животных клетках видны звездообразные фигуры у полюсов. Они образуются вокруг центриолей, разошедшихся к полюсам клетки.
Хромосомы (каждая из двух хроматид) выстраиваются в плоскости экватора, образуя так называемую метафазную пластинку. Длинные хромосомы, как правило, изогнуты V-образно. При этом острие V указывает в центр метафазной пластинки и на нем видна неокрашенная перетяжка – центромера, к которой прикрепляется нить ахроматинового веретена, обеспечивающая движение хромосомы. Центромеры выстраиваются в экваториальной плоскости, а плечи достаточно длинных хромосом словно свисают к тому или другому полюсу.
Анафаза. Это очень короткая стадия. Каждая центромера расщепляется на две, и нити веретена оттягивают дочерние центромеры к противоположным полюсам. Центромеры тянут за собой отделившиеся одна от другой хроматиды, которые теперь называются хромосомами.
Телофаза – заключительная стадия митоза – начинается завершением движения хромосом к полюсам.Хромосомы достигают полюсов клетки, деспирализуются, удлиняются, и их уже нельзя четко различить.
Нити веретена разрушаются, а центриоли реплицируются. Вокруг хромосом на каждом из полюсов образуется ядерная оболочка. Вновь появляется ядрышко. За телофазой может сразу следовать цитокинез (разделение всей клетки на две).
Задание 2.3 Изучить механизмы репродукции и дифференцировки половых клеток.
Мейоз – это два последовательных деления в половых клетках – редукционное и митоз (эквационное или уравнительное деление).
Целью мейоза является образование у животных половых клеток с гаплоидным набором хромосом, что позволяет осуществлять половое размножение, которое является ведущим способом образования новых организмов у многоклеточных животных.



1 2 3



4 5 6


7 8 9
Рисунок 7. Схематическое изображение стадий мейоза:
1 – ранняя профаза, 2 – профаза I, 3 – поздняя метафаза I, 4 – анафаза I, 5 – телофаза I, 6 – профаза II, 7 – метафаза II, 8 – анафаза II, 9 – стадии, следующие за телофазой II, разделение цитоплазмы.
Интерфаза. Происходит репликация органелл, и клетка увеличивается в размерах. Репликация ДНК и гистонов в основном заканчивается в премейотической интерфазе, но частично захватывает и начало профазы I.
Каждая хромосома представлена теперь парой хроматид, соединенных центромерой. Хромосомный материал окрашивается, но из всех структур четко выявляются только ядрышки.
Профаза I. Самая продолжительная фаза. Ее часто делят на пять стадий: лептонема, зигонема, пахинема, диплонема и диакинез.
В раннюю профазу I (рис. 7.1) хромосомы укорачиваются и становятся видимыми как обособленные структуры. У некоторых организмов они выглядят как нитки бус: участки интенсивно окрашивающегося материала – хромомеры – чередуются у них с неокрашивающимися участками.
Затем (рис. 7.2) гомологичные хромосомы, происходящие из ядер материнской и отцовской гамет, приближаются одна к другой и конъюгируют. Эти хромосомы одинаковой длины, их центромеры занимают одинаковое положение, и они обычно содержат одинаковое число генов, расположенных в одной и той же линейной последовательности. Хромомеры гомологичных хромосом лежат бок о бок. Процесс конъюгации называют также синапсисом он может начинаться в нескольких точках хромосом, которые затем соединяются по всей длине.
Пары конъюгировавших гомологичных хромосом часто называют бивалентами. Биваленты укорачиваются и утолщаются. При этом происходит как более плотная упаковка на молекулярном уровне, так и внешне заметное закручивание (спирализация). Теперь каждая хромосома с ее центромерой ясно видна.
Гомологичные хромосомы, составляющие бивалент, частично разделяются, как будто отталкиваясь друг от друга. Теперь видно, что каждая хромосома состоит из двух хроматид.
Хромосомы все еще соединены между собой в нескольких точках. Эти точки называют хиазмами (от греч chiasma -перекрест). В каждой хиазме происходит обмен участками хроматид в результате разрывов и воссоединений, в которых участвуют две из четырех имеющихся в каждой хиазме нитей.
В результате гены из одной хромосомы (например, отцовской – А, В, С) оказываются связанными с генами из другой хромосомы (материнской – а, b, c), что приводит к новым генным комбинациям в образующихся хроматидах. Этот процесс называют кроссинговером.
Гомологичные хромосомы после кроссинговера не расходятся, так как сестринские хроматиды (обеих хромосом) остаются прочно связанными вплоть до анафазы.
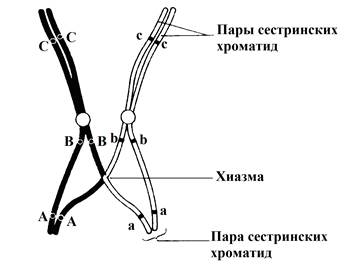
Рисунок 8. Кроссинговер во время профазы I
Хроматиды гомологичных хромосом продолжают отталкиваться друг от друга, и биваленты приобретают определенную конфигурацию в зависимости от числа хиазм.
К концу профазы I все хромосомы полностью уплотнены и интенсивно окрашиваются.
В клетке происходят и другие изменения – миграция центриолей (если они имеются) к полюсам, разрушение ядрышек и ядерной мембраны, а затем образование нитей веретена.
Метафаза I. Биваленты выстраиваются в экваториальной плоскости, образуя метафазную пластинку. Их центромеры ведут себя как единые структуры (хотя часто выглядят двойными) и организуют прикрепленные к ним нити веретена, каждая из которых направлена только к одному из полюсов.
В результате слабого тянущего усилия этих нитей каждый бивалент располагается в области экватора, причем обе его центромеры оказываются на одинаковом расстоянии от него – одна снизу, а другая сверху.
Анафаза I. Имеющиеся у каждого бивалента две центромеры еще не делятся, но сестринские хроматиды уже не примыкают одна к другой. Нити веретена тянут центромеры, каждая из которых связана с двумя хроматидами, к противоположным полюсам веретена. В результате хромосомы разделяются на два гаплоидных набора, попадающих в дочерние клетки.
Телофаза I. Расхождение гомологичных центромер и связанных с ними хроматид к противоположным полюсам означает завершение первого деления мейоза.
Число хромосом в одном наборе стало вдвое меньше, но находящиеся на каждом полюсе хромосомы состоят из двух хроматид. Вследствие кроссинговера при образовании хиазм эти хроматиды генетически не идентичны, и при втором делении мейоза им предстоит разойтись. Веретена и их нити обычно исчезают.
У животных и у некоторых растений хроматиды деспирализуются, вокруг них на каждом полюсе формируется ядерная мембрана и образовавшееся ядро вступает в интерфазу. Затем начинается деление цитоплазмы (у животных) как при митозе.
Интерфаза II. Фаза S отсутствует, и дальнейшей репликации ДНК не происходит. Процессы, участвующие во втором делении мейоза, по своему механизму сходны с происходящими в митозе. Они включают разделение хроматид в обеих дочерних клетках, получившихся в результате первого деления мейоза.
Профаза II. В клетках, у которых выпадает интерфаза II, эта стадия тоже отсутствует. Продолжительность профазы II обратно пропорциональна продолжительности телофазы I.
Ядрышки и ядерные мембраны разрушаются, а хроматиды укорачиваются и утолщаются. Центриоли, если они есть, перемещаются к противоположным полюсам клеток; появляются нити веретена. Хроматиды располагаются таким образом, что их длинные оси перпендикулярны оси веретена первого деления мейоза.
Метафаза II. При втором делении центромеры ведут себя как двойные структуры. Они организуют нити веретена, направленные к обоим полюсам, и таким образом выстраиваются по экватору веретена.
Анафаза II. Центромеры делятся, и нити веретена растаскивают их к противоположным полюсам. Центромеры тянут за собой отделившиеся друг от друга хроматиды, которые теперь называются хромосомами.
Телофаза II. Эта стадия очень сходна с телофазой митоза. Хромосомы деспирализуются, растягиваются и после этого плохо различимы. Нити веретена исчезают, а центриоли реплицируются. Вокруг каждого ядра, которое содержит теперь половинное (гаплоидное) число хромосом исходной родительской клетки, вновь образуется ядерная мембрана.
В результате последующего деления цитоплазмы из одной исходной родительской клетки получается четыре дочерних клетки.
Раздел 2
МОЛЕКУЛЯРНЫЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
тема 3
Строение и синтез нуклеиновых кислот
Нуклеиновые кислоты – сложные биологические полимеры, состоящие из более простых соединений (нуклеотидов).
ДНК – уникальное химическое соединение, не имеющее аналогов – она способна к самовоспроизведению (авторепродукции). Благодаря чередованию четырех азотистых оснований молекулы ДНК бесконечно разнообразны.
| Компоненты | ДНК | РНК |
| Пуриновые основания | Аденин, гуанин | Аденин, гуанин |
| Пиримидиновые основания | Цитозин, тимин | Цитозин, урацил |
| Углеводный компонент | Дезоксирибоза | Рибоза |
Нуклеотиды ДНК состоят из остатка фосфорной кислоты (Ф), сахара – дезоксирибозы (Д) и одного из четырех азотистых оснований: пуриновых – аденина (А), гуанина (Г); пиримидиновых – тимина (Т), цитозина (Ц).
Нуклеотиды ДНК:
Ф — Д Ф — Д Ф — Д Ф — Д
 | | | |||||
 | |||||||
А Г Т Ц
адениновый гуаниновый тиминовый цитозиновый
Молекула ДНК состоит из двух связанных между собой полинуклеотидных цепей, скрученных в виде спирали. Для записи расположения нуклеотидов во второй цепи ДНК, надо знать правило комплементарности азотистых оснований: аденин всегда связан с тимином, а гуанин – с цитозином. Эти пары азотистых оснований комплементарны по отношению друг к другу, то есть дополняют друг друга.
Нуклеотиды в молекулах ДНК и РНК связываются между собой через фосфорную кислоту. Аденин и тимин соединены двойными водородными связями, гуанин с цитозином – тройными.
В 1950 г. Э. Чаргафф установил правило нуклеотидных отношений, лежащее в основе строения всех ДНК.
Правило Чаргаффа состоит в том, что в ДНК содержание аденина равно содержанию тимина (А=Т), а содержание гуанина равно содержанию цитозина (Г=Ц), отсюда А+Г/Т+Ц=1; сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклеотидов. В соответствии с этим правилом нуклеотидный состав разных организмов может варьировать только по величине А+Т/Г+Ц.
Расстояние между нуклеотидами 3,4 Å (1 Å – ангстрем равен десятимиллионной доле миллиметра) или 0,34 нм – нанометра. Длина пуриновых оснований равна 12 Å, пиримидиновых оснований – 8 Å. Диаметр двойной цепи 20 Å.
Молекула ДНК способна к авторепродукции (самовоспроизведению), при которой она воспроизводит две молекулы с той же самой уникальной последовательностью пар азотистых оснований; авторепродукция происходит в интерфазе митоза.
Спираль родительской ДНК раскручивается, и комплементарные цепи отделяются одна от другой в результате разрыва слабых водородных связей между комплементарными основаниями (рис. 9а, б).

Рисунок 9. Авторепродукция ДНК
Происходит спаривание между основаниями матричной цепи и свободными нуклеозидтрифосфатами, последние связываются друг с другом, образуя новую полинуклеотидную цепь (рис. 9в). Таким образом, получаются две идентичные молекулы ДНК.
Молекула РНК представляет собой длинную цепь, состоящую из последовательно расположенных звеньев – нуклеотидов. Вещества с таким строением называют линейными полимерами, а входящие в состав цепи звенья – мономерами.
В отличие от ДНК, РНКявляется, в основном, одноцепочной и неспособна у многоклеточных организмов к делению и к самовоспроизведению. Исключение составляют вирусы, которые синтезируются на молекулах ДНК.
Нуклеотид состоит из трех компонентов – остатка фосфорной кислоты (фосфата), пятиатомного сахара – рибозы (Р) и одного из четырех азотистых оснований: аденина (А), гуанина (Г), цитозина (Ц) и урацила (У).
Нуклеотиды РНК:
Ф — Р Ф — Р Ф — Р Ф — Р
| | |
А Г У Ц
адениновый гуаниновый урациловый цитозиновый
Молекула РНК состоит из остова (регулярно чередующихся фосфата и рибозы) и ответвляющихся от остова азотистых оснований (А, Г, Ц, У).
Не нашли, что искали? Воспользуйтесь поиском: