ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Уплыў на фотасінтэз іншых экалагічных фактараў.
Роля цяпла. Цяпло – вымяраецца тэмпературай. Ад цяпла меньш залежаць рэакцыі светлавой фазы (фотахімічныя рэакцыі) і цалкам залежаць ферментатыўныя рэакцыі цемнавой фазы. Пры павышэнні тэмпературы да аптымальных значэнняў на кожныя 10°С інтэнсіўнасць фотасінтэзу прыкладна падвойваецца (Q10 = 2-3). Пры больш высокіх тэмпературах інактывіруюцца ферменты і фотасінтэз зніжаецца да нуля.
Крывая залежнасці фотасінтэзу ад цяпла (тэмпературная крывая) мае тры кардынальныя кропкі: мін, оптымум і макс. Значэнні кардынальных кропак – гэта ўласцівасці, набытыя пры адаптацыі раслін ў філагенезе да ўмоў росту. Яны вызначаюцца геаграфічным паходжаннем раслін. Чым больш на поўдзень знаходзіцца арэал віда, тым вышэй гэтыя паказчыкі.

У раслін умеранага клімату фотасінтэз пачына-ецца пры адмоўных тэмпературах -5-7°С – ніжняя граніца замярзання тканак. Найбольшых значэнняў чысты фотасінтэз дасягае пры 20-25°С (оптымум), сапраўдны – пры 30-35°С. Максімальныя значэнні тэмператур, пры якіх сапрадны фотасінтэз спыняецца – 40-45°С. Тэмпературны кампенсацыйны пункт, калі брута фотасінтэз роўны дыханню, а чысты роўны нулю, назіраецца пры 35-40°С.
Для параўнання у субтрапічных раслін міні-мальныя тэмпературы для фотасінтэзу +5-7°С, оптымум – 30-35°С, максімальныя – 50-55°С. Да гэтай групы раслін адносяцца таматы, агуркі, многія кветкавыя расліны.
На фотасінтэз адмоўна ўплываюць рэзкія зрухі тэмпературы, наступствы якіх праяўляюцца і пры нармалізацыі ўмоў. Пры ўздзеянні ў доследах на сеянцы хваёвых раслін адмоўнай тэмпературай летам фотасінтэз аднаўляўся толькі праз некалькі дзён. Чым меньш былі тэмпературы, тым больш часу праходзіць да аднаўлення фотасінтэзу. Пры выпрабаванні тэмпературай -10°С ні адзін з відаў не аднавіў фотасінтэз. Інгібіруюць фотасінтэз таксама і высокія тэмпературы. Тэмпературны дыяпазон фотасінтэзу можна расшырыць закальваннем расліны нізкімі і высокімі тэмпературамі.
Роля СО2. Залежнасць фотасінтэзу ад канцэнтрацыі СО2 мае форму лагарыфмічнай крывой. Хуткі рост фотасінтэзу назіраецца ад 0 да 0,03%. Найбольшых значэнняў фотасінтэз дасягае пры павелічэнні ўтрымання вуглекіслаты ў паветры да 0,3% – на парадак больш, чым у звычайных умовах, а пры вялікай асветленасці – да 1,5-2%. Пры канцэнтрацыях СО2 большых за аптымальныя закрываюцца вусцейкі, закісляецца змесціва клетак. Змяненні рН прыводзяць да зніжэння актыўнасці ферментаў і парушэнняў абмену рэчываў.
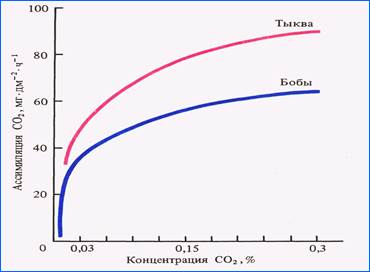
Колькасць СО2 у паветры ўтрымліваецца на пастаянным узроўні. Паглынанне яго раслінамі кампенсуецца дыханнем жывых арганізмаў, асабліва глебавых, разлажэннем арганікі глебы. У лясным фітацэнозе колькасць СО2 вагаецца на працягу сутак прыкладна на 25 % ад сярэдняга ўзроўню. Назіраецца таксама вертыкальны градыент канцэнтрацыі вуглякіслаты: у прыземным паветры яе больш, у кронах – меньш.
У цяпліцах расліны могуць галадаць з-за недахопу вуглякіслаты, таму яе колькасць павышаюць да 1-2%. Адначасова забяспечваюцца неабходныя ўмовы па цяплу і асветленасці. Гэта дазваляе павысіць ураджай да 30%.
Павелічэнню колькасці карбанату каля расліны садзейнічае ўнясенне арганікі і рыхленне глебы.
Роля вільготнасці глебы. Ад вільготнасці залежыць водны баланс расліны. Недахоп вады прыводзіць да зніжэння воднага патэнцыялу ліста і павелічэння воднага дэфіцыту, закрыцця вусцейкаў і ў выніку вуглекіслотнага галадання расліны.
Пры паніжэнні воднага патэнцыялу фотасінтэз падаўляецца хутчэй, чым дыханне, таму расліна пачынае расходаваць арганічных рэчываў больш, чым сінтэзаваць.

Водны дэфіцыт ўплывае на святлохімічныя і біяхімічныя працэсы: вада з’яўляецца крыніцай электронаў і пратонаў для фотасінтэзу; пры недахопе парушаецца перанос электронаў па ЭТЛ, фотафасфарыляванне, дзейнасць ферментаў. Шкодны ўплыў тым большы, чым большым было абязводжанне і яго працягласць.
Для кожнай расліны ёсць свой паказчык аптымальнай вільготнасці глебы, сярэдні ж паказчык роўны каля 60 – 70 % яе поўнай вільгацеёмістасці.
Роля мінеральнага жыўлення. Мінеральныя элементы ўваходзіць у склад арганічных рэчываў. Яны неабходны для росту лісця, утварэння ферментаў, якія каталізуюць працэсы фотасінтэзу, рэгуляцыі дзейнасці ферментаў. Так, Mg2+ уваходзіць у склад хларафілу і з’яўляецца кафактарам фермента, які каталізуе працэс карбаксілявання РДФ (рыбулёза дыфасфату); іёны Fe2+ неабходны для сінтэзу хларафілу, яны ўваходзяць у склад цытахрома, які пераносіць электроны ў дыхальным ланцугу; азот таксама ўваходзіць у склад хларафілу; калій неабходны для падтрымання структуры хларапластаў, удзельнічае ў працэсе адкрывання і закрывання вусцейкаў.
Пры недахопе Mg, Fe, N у расліны адбываецца парушэнне абмену рэчываў, у выніку чаго памяньшаецца колькасць хларафілу і лісце набывае жоўта-зялёную афарбоўку. Гэтая з’ява называецца хлароз. Хлароз можа назірацца і пры ўзнікненні іншых неспрыяльных умоў, захворванні расліны.
Роля кіслароду. Кісларод уплывае на расліну праз дыханне. Энергія дыхання неабходна для працэсаў сінтэзу. Мінімальная колькасць кіслароду, якая патрэбна расліне – 5% (у паветры 21%).
Кісларод у С3-раслін стымулюе фотадыханне, якое адбываецца толькі ў зялёных частках расліны на святле (яно адрозніваецца ад звычайнага дыхання). У працэсе фотадыхання расходуюцца прадукты фотасінтэзу.Чым большая канцэнтрацыя кіслароду, тым большае фотадыханне і меньшы рэзультат фотасінтэзу. У С4-раслін фотадыханне адсутнічае і яны маюць большую прадукцыйнасць.
Сумеснае дзеянне фактараў.
На расліну дзейнічае комплекс фактараў. Інтэнсіўнасць фотасінтэзу вызначаецца тым фактарам, які знаходзіцца ў адносным мінімуме (закон мінімума Ю.Лібіха).

З графіка відаць, што пры асветленасці 100 Дж*см2*сек тэмпература 12ºС абмяжоўвае фотасінтэз, а пры тэмпературы 31ºС абмежавальным фактарам з’яўляецца святло. Светлавое насычэнне назіраецца пры асветленасці ў 200 адзінак.
Тое ж адбываецца і пры сумесным уздзеянні на фотасінтэз святла і вуглекіслаты.

Часцей за ўсё фотасінтэз абмяжоўваецца наступнымі фактарамі: у цёплыя ясныя дні – недахопам СО2; у пачатку і канцы дня і пад полагам лесу – недахопам святла; таксама нізкай і высокай тэмпературай; недахопам вады ў перыяд засухі, асабліва на ўзвышшах.
Максімальны фотасінтэз дасягаецца пры аптымізацыі ўсіх фактараў.
Сутачная і сезонная дынаміка фотасінтэзу раслін. На працягу сутак пры спрыяльных умовах вільготнасці фотасінтэз расце да сярэдзіны дня, потым зніжаецца. Пры недахопе вады крывая 2-х вяршынная – удзень зачыняюцца вусцейкі. Пры хранічным недахопе пік фотасінтэзу назіраецца толькі раніцою, вусцейкі застаюцца зачыненымі і ў другой палове дня. Многавершынная крывая назіраецца пры пераменнай воблачнасці.

У лістападных раслін фотасінтэз расце вясною з развіццём лісця і падае увосень. У хваёвых пачынаецца рана вясною з наступленнем дадатных тэмператур і заканчваецца увосень з прыходам марозу. Працягласць перыяду, пры якім адбываецца фотасінтэз у хваёвых, большы, чым гэты перыяд у лісцевых. Хваёвыя расліны меньш расходуюць арганічных рэчываў на дыханне. Дзякуючы гэтаму, хваёвыя расліны фарміруюць такія ж па прадукцыйнасці дрэвастоі, што і лісцевыя, хаця інтэнсіўнасць фотасінтэзу ў іх значна меньшая. Так, Iф ёлкі прыкладна ў 5 разоў меньшая, чым у бярозы.
На самастойную работу.
Не нашли, что искали? Воспользуйтесь поиском: