ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
ВЗАИМОДЕЙСТВИЕ ГЕНОВ: ТИПЫ ВЗАИМОДЕЙСТВИЙ, БИОХИМИЧЕСКИЕ ОСНОВЫ.
I.Взаимодействие аллельных генов: 1.Доминирование – признаки, контролируемые геном в аутосоме перед-ся по аутосомно-доминантн.типу (равновероятно передаются и ♀, и ♂ потомкам), признаки, контролируемые геном в пол.хромосоме – по сцепленному с полом доминант.типу (перед-ся от отца только дочерям). Некоторые дом.мутации в гомозиг.состоянии – летальные; 2.Неполн.доминирование – как у кур андалузской породы (при скрещивании гомозигот с белым и черным оперением в F1 получают серых кур); 3.Кодоминирование – проявляются оба аллеля у гетерозигот (i.e., наледование групп крови у человека - три аллеля домин. IAи IB, рецесс. I0. IAIA или IAI0 – группа А, I0I0 – группа 0, IAIB – группа АВ, т.е.проявляются оба аллеля =>имеются оба типа поверх.антигенов); 4.Сверхдоминирование, гетерозис – усиление признака у гетерозигот (i.e.большая плодовитость у гетерозиготных мух, чем у исх.форм); 5 .Неустойчивая доминантность – проявление признака у гетерозигот зависит от внешних условий и генотип.окружения (i.e., доминантная мутация Curly не проявляется в форме фенотипа с загнутыми вверх крыльями при 19°С; доминант.аллель w+ в рез-те инверсии попадает в прицентромерн.хроматин =>у гетерозигот w+/w проявляется рецесс.аллель w – белые глаза). 6.Условная доминантность – невозможность выявить гомозигот по домин.аллелю, т.к.такие особи нежизнеспособны (доминант.мутация действует летальна в гомозиготе).
II.Взаимодействие аллельных генов: 1.Комплементарность – два гена «работают» вместе =>развитие отличного от родит.варианта признака. Три типа: дом.гены разл-ся по фенотип.проявлениям, дом.гены имеют сходное проявление, и дом. и рец. гены имеют самостоят.фенотип.проявление. Примеры: ¶ наследование формы гребня у кур – A_B_ имеют ореховид.форму, A_bb – розовидную, bbA_ - гороховидную, aabb – обычную; ¶ наследование окраски кокона у тутового шелкопряда – желтые коконы только у A_B_, при наличии только одного дом.гена и двойных гомозигот по реццессиву – неокрашенные коконы. 2.Эпистаз. а.Доминантный эпистаз – дом.ген подавляет проявление другого дом.гена (i.e., у тыквы желтую окраску плода опред-ет ген А, зеленую опред-ет а, в присутствии дом.ингибитора I – окраски нет, I_A_ и I_aa имеют бесцветные плоды). б.Рецессивный эпистаз – рец.аллель одного гена подавляет, а между доминантными генами наблюд-ся комплементарность. При двойном эпистазе каждая гомозиг.рецесс.аллель подавляет домин.аллель другого гена. 3.Полимерия – гены дублируют действие друг друга. Два типа: некумулятивная (i.e.образование овального стручка только у пастушей сумки с генотипом a1a1a2a2, и треугольного – у всех остальных) и кумулятивная (окраска зерен пшеницы пропорциональна числу доминантных генов, самая интенсивная – у A1A1A2A2A3A3).
Комплементарный тип взаимодействия (взаимно дополнительный) осуществл., когда доминантные аллели обоих генов (в дигибридном скрещивании) обусловливают нормальный (или дикий) фенотип.
Пример: PP st st bw+ bw+ × st+ st+ bw bw (ярко-красные×коричневые глаза)
F1 st+_ bw+_ (темно-красные глаза)
F2 9 st+_ bw+_ (темно-красные глаза): 3 st+_ bw bw (коричневые глаза): 3 st st bw+_ (ярко-красные глаза):1st st bw bw (белые глаза)
Темно-красные глаза- дикий тип; ярко-кр. и кор. глаза рецессивны по отнош. к дикому типу, бел.- двойной рецессив.
Биохим. мех-м взаимод. аллелей st и bw исслед. подробно.У дрозофилы окраска глаз обусловлена синтезом 2-х пигментов - кр. и кор. Рецессив bw в гомозиготе прерывает синтез кр. пигмента; рецессив st - кор. Когда в дигетерозиготе оказыв. норм. аллели, синтез. оба пигмента. Если в гомозиготе оказыв. и bw bw, и st st, то не синтез. ни кр., ни кор. пигменты, и глаза - белые.
По типу комплементарности взаимод. гены, контрол. разные этапы одного и того же метабол. пути,однако для многих неизвестен биохим. мех-м их реализации. По типу комплементарности взаимод. гены, определ. форму гребня кур, форму плода у тыквы.
Эпистаз- рецессивная аллель в гомозиготе препятствует проявлению доминантной аллели или, наоборот, доминантная аллель препятствует проявлению рецессивной аллели. Пример: синтез кор. пигмента у мух, кроме st, блокируют также и рецессивные аллели гена purple(pr). Выход в гомозиготу любой из 2 рецессивных аллелей блокирует синтез пигмента; появлениеи st, и pr не приводит к белым глазам(ярко-кр.).Это тоже пример комплементарного взаимод., но без новообразования. В данном случае рецессивная аллель pr эпистатична по отношению к доминантной аллели st+, а st эпистатична по отношению к pr+(двойной рецессивный эпистаз). Существ. также простой рецессивный эпистаз (a > B; a > b или b >A; b>a), кот. выраж. в расщеплении 9:3:4; простой доминантный эпистаз (A >B;A>b или B>A; B>a) с расщеплением 12:3:1. Ген, подавляющий действие другого, назыв. ингибитором или супрессором; подавляемый ген - гипостатический.
Полимерия - одной доминантной аллели любого из взаимод. генов достаточно для проявления фенотипа. Пример: пастушья сумка PP треуг. стручки×овал. стручки
F1 все треуг. стручки
F2 15 треуг.: 1 овал.(а1а1а2а2) Гены в этом случае обознач. Одинаково (А1 и А2). Это некумулятивная полимерия.
Кумулятивная полимерия (Нильсон-Эле, 1908) - интенсивность проявления признака зависит от числа доминантных аллелей (при тригибридном скрещивании расщепление 1:6:15:20:15:6:1). Так наследуются окраска эндосперма зерен пшеницы, цвет кожи у человека, молочность, яйценоскость, длина колоса у злаков, содержание сахара в корнеплодах сахарной свеклы. Изучением наследования таких признаков занимается генетика количественных признаков.
13. СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ И КРОССИНГОВЕР
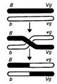 [На примере наследования признака окраски тела (b – рец., черное тело/ b+ - дом., серое тело) и типа крыльев (vg – рец., редуцир.клетки/ vg+ − норм.крылья.). 1.Полное сцепление (Дигетерозиготный самец и гомозиготная по рецессивам самка): P ♂ b vg+/b+ vg × ♀ b vg/b vg =>
[На примере наследования признака окраски тела (b – рец., черное тело/ b+ - дом., серое тело) и типа крыльев (vg – рец., редуцир.клетки/ vg+ − норм.крылья.). 1.Полное сцепление (Дигетерозиготный самец и гомозиготная по рецессивам самка): P ♂ b vg+/b+ vg × ♀ b vg/b vg =>
=> F1 50% b+vg/b vg и 50% b vg+/b vg => самцы образуют только два типа гамет (b vg+ и b+ vg). 2.Неполное сцепление (Дигетерозиготная самка и гомозиготный по рецессивам самец). Р ♀ b vg+/b+ vg × ♂ b vg/b vg => 83% - b vg+/b vg и b+vg (есть сцепление генов) и 17% b vg/b vg и b+vg+/b vg (рекомбинантные формы, сцепления нет) =>полное сцепление имеется только у самцов. 3.Кроссинговер. Морган объяснил неполное сцепление обменом участками между генами b и vg. (см.также вопрос 15).
14. Интерфеенция – подавление кроссинговера на участках, непосредственно прилегающих к точке произошедшего обмена. Коэффициент коинциденции С=налюдаемая частота двойных кроссинговеров/теоретич ожидаемая. С<1=> положительная интерференция, C>1=> отрицательная.
В дальнейшем Т. X. Морган и его сотрудники в экспериментах с D. melanogaster обнаружили большое число примеров сцепления генов и показали, что это сцепление, как правило, неполное. Рассмотрим один из первых экспериментов Т. X. Моргана по изучению сцепленного наследования. У дрозофилы известны мутантные формы, отличающиеся от мух дикого типа черной окраской тела. Это признак рецессивный по отношению к признаку нормальной cерой окраски. Ген, контролирующий черную окраску тела, называется black и обозначается b. Его доминантная аллель — b+. Существует также рецессивный ген vestigial (vg), который в гомозиготном состоянии приводит к недоразвитию крыльев (зачаточные крылья). Его доминантная аллель (vg+) контролирует нормальное развитие крыльев.
При скрещивании мух bb vg vg X b+ b+ vg+ vg+ в F1 были получены особи, дигетерозиготные по этим генам. Все они были нормальными по обоим признакам в соответствии с правилом доминирования и законом единообразия F1. Далее были проведены два типа анализирующих скрещиваний. В первом из них брали самцов F1 и скрещивали с гомозиготными самками bbvg vg, а во втором — девственных самок, отобранных в F1, скрещивали с самцами bbvgvg. Результаты этих анализирующих скрещиваний оказались неодинаковыми (рис. 5.10, А). В Fа (потомство от анализирующего скрещивания) в первом случае были получены мухи только двух типов независимо от пола: 50 % мух имели черное тело и зачаточные крылья и 50 % были нормальными по обоим признакам. Учитывая, что расщепление в анализирующем скрещивании отражает соотношение типов гамет, продуцируемых особями fi, следует заключить, что самцы F1, использованные при первом скрещивании, формировали гаметы только двух типов — с родительскими сочетаниями аллелей bvg и b+ vg+. Следовательнл в 100% случаев были гаметы только с родительскими сочетаниями исследов. генов.При втором скрещивании в Fa появились все возможные четыре типа потомков, а следовательно, самки fi давали четыре типа гамет: b vg, b+ vg, b vg+ и b+ vg+. Однако, как явствовало из расщепления в Fa от этого скрещивания, четыре типа гамет образовались не равновероятно. Независимо от пола мухи распределились следующим образом: 41,5 % черных с зачаточными крыльями; 41,5 % нормальных по окраске и с нормальными крыльями; 8,5 % черных с нормальными крыльями; 8,5 % нормальных по окраске с зачаточными крыльями. Таким образом, родительские сочетания b vg и b+ vg+ образовались в 83 % случаев, а новые комбинации — рекомбинантные сочетания bvg+ и b+vg (рис. 5.10, А) — в 17 % случаев.
В другом эксперименте в качестве родителей были использованы мухи, обладающие теми же признаками, но в другом сочетании: мух с черным телом и нормальными крыльями скрещивали с мухами с нормальной окраской тела и с зачаточными крыльями. Затем вновь провели два типа анализирующих скрещиваний. В первом использовали самцов fi, во втором — самок F1. В обоих случаях их скрещивали с двойным гомозиготным рецессивом bbvg vg. И вновь были получены такие же результаты в отношении родительских и рекомбинантных сочетаний признаков в Fа. Если из F1 брали самцов, то наблюдали только
родительские комбинации признаков, а если из fi брали самок, то появлялись родительские (83%) и рекомбинантные (17%) сочетания признаков в тех же соотношениях, что и в первом эксперименте, результаты которого представлены на рис. 5.10, А.
В обоих экспериментах наблюдается полное сцепление генов b и vg, если для анализирующего скрещивания берутся самцы fi. Если же для анализирующего скрещивания использовали самок fi, то сцепление было частичным.
Т. X. Морган дал следующее объяснение этим результатам. В первом эксперименте гены b и vg находятся в одной хромосоме, т. е. дигетерозиготные особи fi несут в одном гомологе аллели b и vg, а в другом гомологе — b+ и vg+ (рис. 5.10, А); во втором эксперименте b и vg+ в одном, а b+ и vg(рис. 5.10, Б).
У самцов дрозофилы кроссинговер вообще не происходит, поэтому гены, локализованные в одной хромосоме (или, говоря более строго, в одной паре хромосом), обнаруживают абсолютное сцепление, если при скрещивании используют дигетерозиготных самцов. В мейозе у дигетерозиготных самок дрозофилы fi возможен обмен гомологичными участками гомологичных хромосом между локусами, в которых находятся гены b и vg. Такие обмены, или кроссинговер (от англ, crossingover — перекрест), приводят к новому рекомбинантному сочетанию аллелей генов b и vg в гомологичных хромосомах, которые затем расходятся к разным полюсам. Эти обмены происходят с вероятностью 17 % ив итоге дают два класса реципрокных рекомбинантных сочетаний, или ре-комбинантов с равной вероятностью — по 8,5 % (рис. 5.11).
Сходным образом объясняется и результат, полученный ранее У. Бэтсоном и Р. Пеннетом: гены, контролирующие окраску цветков (р) и форму пыльцевого зерна (/) у душистого горошка, локализованы в одной паре гомологичных хромосом, и между ними возможен кроссинговер. Сотрудник Т. X. Моргана А. Стертевант предположил, что частота кроссинговера на участке между генами, локализованными в одной хромосоме, может служить мерой расстояния, на котором они находятся друг от друга. Тогда можно использовать частоту кроссинговера для того, чтобы определять взаимное расположение генов и расстояние между ними.
В качестве подтверждения справедливости этого положения можно в общем виде рассмотреть результаты тригибрибридного скоещивания,в кот. Род. Формы дрозофилы различаются по генам b и vg, pr(сцеплен. c b и vg).
Рецессивная аллель pr (purple — пурпурный) в гомозиготном состоянии обусловливает ярко-красную окраску глаз, нормальная доминантная аллель рr+ — темно-красный цвет глаз.
При анализирующем скрещивании потомки расщепляются на 8 классов,2 класса нерекомбинантных (/ и //) и 6 классов потомков, рекомбинантных по всем генам (Ш — VIII).
Далее необходимо определить частоту кроссинговера между всеми тремя генами попарно. Для этого суммируют всех мух, рекомбинантных по генам b и рr. классы III, IV, VII, VIII). Полученное число делят на общее число исследованных потомков в Fa. Аналогично определяют частоту рекомбинации (кроссинговера) между рr и vg (при этом суммируют классы V, VI, VII, VIII) и частоту рекомбинации между b и vg (суммируют классы III, IV, V, VI).
Экспериментально установленные частоты рекомбинации между тремя генами попарно можно представить следующим образом: b - рr — 6 %; рr - vg - 12 %; b - vg - 17 %.На основе этих данных, пользуясь - правилом аддитивности, можно расположить три гена в линейной последовательности. Наиболее удалены друг от друга гены b и vg, а между ними локализован ген рr. Сумма час тот его рекомбинации с генами b и vg приблизительно равна частоте рекомбинации между b и vg. Такимобразом, строится простейшая карта группы сцепления. Под группой сцепления понимают группу генов, расположенных в одной хромосоме.
даже гены, расположенные в одной хромосоме, не всегда обнаруживают сцепление. Генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %, представляет собой сантиморган (сМ) — единицу измерения, названную в честь Т. X. Моргана
Интерференция
Так же, как в рассмотренном случае, сумма меньших частот рекомбинации (генетических расстояний) чаще всего превышает частоту рекомбинации между наиболее удаленными друг от друга маркерами. Это объясняется тем, что между любыми двумя сцепленными генами возможен не только одиночный, но и двойной (а также множественный) кроссинговер, что приводит к сокращению регистрируемой частоты кроссинговера. Действительно, если бы в рассмотренном примере (рис. 5.12) между генами b и vg не было бы маркера рr, то b (pr+) vg и b+ (pr) vg+ воспринимались бы как некроссоверные состояния b vg и b+ vg+. Таким образом, двойные обмены сокращают регистрируемое расстояние между генами. Вместе с тем между обменами на соседних участках хромосом существует взаимовлияние, названное интерференцией. Такое взаимовлияние можно выразить количественно. Для этого сопоставляют реально наблюдаемую частоту двойных обменов с частотой, теоретически ожидаемой на основе предположения о том, что обмены на соседних участках происходят независимо друг от друга. Степень и характер интерференции измеряется величиной коинци-денции (С). Коинциденцию оценивают как частное от деления реально наблюдаемой частоты двойных кроссоверов на теоретически ожидаемую частоту двойных кроссоверов. Последнюю величину получают, перемножая частоты кроссинговера на соседних участках.
Вычислим коинциденцию на конкретном примере, пользуясь данными Т. X. Моргана и А. Стертаванта, которые при тригибридном скрещивании изучали-рекомбинацию между генами у, w и m,
локализованными в Х-хромосоме D. melanogaster. Фенотипическое проявление генов у и w уже описывалось. Рецессивная аллель гена m приводит к уменьшению размера крыльев. Частота рекомбинации между у и w 1,3 %, а между w и т 32,6 %. Двойные рекомбинанты по у — w — т наблюдались с частотой 0,045 %.
C(коинциденция)=двойные кроссоверы/произведение недвойн.кроссоверов Величину интерференции (/) определяют по формуле I= 1 — С.
Если С < 1, то интерференция положительная, т. е. одиночный обмен препятствует обмену на соседнем участке хромосомы. Если С>1, то интерференция отрицательная, т.е. один обмен как бы стимулирует дополнительные обмены на соседних участках. В действительности существует только положительная интерференция при реципрокной рекомбинации — кроссинговере, а кажущееся неслучайным совпадение двух и более обменов, характерное для очень коротких расстояний, — результат нереципрокных событий при рекомбинации
Таким образом, при картировании генов в группах сцепления на основе изучения частот рекомбинации необходимо учитывать две противоположные тенденции. Двойные обмены «сокращают» расстояния между генами, а интерференция препятствует множественным обменам, вероятность которых увеличивается с расстоянием. Как показал для дро-зофилы Г. Меллер, на больших расстояниях (около 35% рекомбинации) интерференция исчезает. Следовательно, наиболее точные данные о частоте кроссинговера можно получить только на достаточно коротких расстояниях — приблизительно до 10 сМ.
В обобщенном виде зависимость частоты рекомбинации от реального расстояния с учетом множественных обменов описывает функция Дж. Хол-дэйна: rf(d)=0.5(1-e-2d)
где rf — картирующая функция (в нашем случае — это частота учитываемых кроссинговеров), d — реальное расстояние, на котором происходят обмены, е — основание натурального логарифма. Функция Холдэйна показывает, что с увеличением расстояния rf приближается к 0,5. Реально это означает, что между генами, расположенными далеко друг от друга, выявляется около 50 единиц рекомбинации. Такую же частоту рекомбинации демонстрируют и гены, находящиеся в разных хромосомах. практически невозможно уловить сцепления между столь удаленными друг от друга генами. Эти гены, хотя и сцеплены физически, находясь в одной хромосоме, будут наследоваться независимо.
.Как теперь хорошо известно, некоторые гены, контролирующие 7 признаков гороха, исследованные Г. Менделем, сцеплены, однако расположены на большом расстоянии друг от друга. В частности, гены а (окраска цветков и семенной кожуры) и i(окраска семян) принадлежат к одной и той же группе сцепления, но расстояние между ними около 200 сМ. В опытах Менделя эти гены наследовались независимо. При скрещивании: AAII X aaii в F2 было получено расщепление 357 А — I —: 132 А — ii: 116 ааI —::34aaii, которое хорошо соответствует теоретически ожидаемому при независимом наследовании (360:120:120:40; X2 = 2,258; р>0,05) (см. гл. 2.2). стояния rf приближается к 0,5.
Вопрос 14
Между любыми двумя сцепленными генами возможен не только одиночный, но и двойной (или множественный) кроссинговер, что приводит к сокращению регистрируемой частоты кроссинговера. Кроме того, между обменами на соседних участках хромосом существует взаимовлияние – интерференция (I), степень и характер которой измеряется величиной коинциденции (С). Коинциденция – частное от деления реально наблюдаемой частоты двойных кроссинговеров на теоретически ожидаемую частоту (если обмены на соседних участках происходят независимо друг от друга). I=1-C. Если C<1, то интерференция положительная, то есть одиночный обмен препятствует обмену на соседнем участке хромосомы. Если C>1, то интерференция отрицательная, то есть один обмен как бы стимулирует дополнительные обмены на соседних участках. В действительности при кроссинговере существует только положительная интерференция.
Не нашли, что искали? Воспользуйтесь поиском: