ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
КАТАБОЛИЗМ УГЛЕВОДОВ
Основные пути катаболизма углеводов - анаэробный и аэробный. Анаэробный путь катаболизма осуществляется клеткой без доступа кислорода воздуха и называется брожением; аэробный - идёт с участием кислорода воздуха, представляет собой окислительный процесс и называется дыханием. Конечные пути обоих путей: а) синтез АТР за счёт утилизации энергии аккумулированной в углеводах; б) получение из этих углеводов пластических соединений, являющихся исходным материалом для синтеза компонентов клетки.
Гликолиз (от греч. глицис - сладкий, лизис - разрушение) - универсальный и основной процесс катаболизма углеводов для большинства организмов, это последовательность реакций, приводящих к превращению гексозы (глюкозы, фруктозы и др.) в пируват с одновременным образованием АТР.
Исследования химизма гликолиза показали, что начальные этапы процесса брожения и дыхания имеют общий путь. При дыхании у аэробных организмов гликолиз предшествует циклу трикарбоновых кислот и цепи переноса электронов. Субстрат переносится в митохондрии, где он полностью окисляется до углекислого газа, в результате чего с высокой эффективностью из гексозы извлекается свободная энергия. При брожении в анаэробных условиях пируват превращается в продукты брожения - лактат и этанол.
Все реакции процесса гликолиза протекают в цитоплазме и катализируются десятью различными ферментами. Эти ферменты растворимы в воде, легко экстрагируются из клеток.
Первая стадия гликолиза начинается с фосфорилирования гексоз и является пусковой. Она требует затраты химической энергии в виде АТР. Пусковая стадия включает: 1) фосфорилирование глюкозы или других гексоз; 2) их изомеризацию; 3) второе фосфорилирование; 4) расщепление на триозофосфаты.
1 реакция. Осуществляется реакция между D-глюкозой и АТР, приводящая к образованию глюкозо-6-фосфата. Реакция катализируется гексокиназой. В различных клетках имеются несколько изоформ гексокиназ, некоторые из них относительно неспецифичны и осуществляют также фосфорилирование других гексоз. В печени имеется глюкокиназа, отличающаяся от гексокиназы значительно меньшим сродством к глюкозе.
2 реакция. Осуществляется изомеризация гексозо-6-фосфатов во фруктозо-6-фосфат - все гексозы превращаются в один фосфорилированный продукт. Реакция катализируется фосфоглюкоизомеразой.
3 реакция. За изомеризацией следует второе фосфорилирование. Под действием фосфофруктокиназы и при участии АТР фруктозо-6-фосфат превращается в фруктозо-1,6-бифосфат.
Почему метаболизм гексоз начинается с двух этапов фосфорилирования? Одна из причин - с кинетической точки зрения длинную последовательность реакций выгодно начинать с практически необратимой стадии - фосфорилирования. Образование фруктозо-1,6-бифосфата является наиболее медленной текущей стадией гликолиза. Она в целом и определяет скорость всего процесса гликолиза.
4 реакция. Далее следует превращение фруктозо-1,6-бифосфата в триозофосфаты - глицеральдегидтрифосфат и дигидроксиацетонфосфат. Эту реакцию катализирует альдолаза. После расщепления фруктозо-1,6-бифосфата в каждой из образовавшихся половин оказывается фосфатная группа.
5 реакция. Под действием триозофосфатизомеразы происходит взаимопревращение фосфотриоз. Равновесие этой реакции сдвинуто в сторону дигидроксиацетонфосфата.
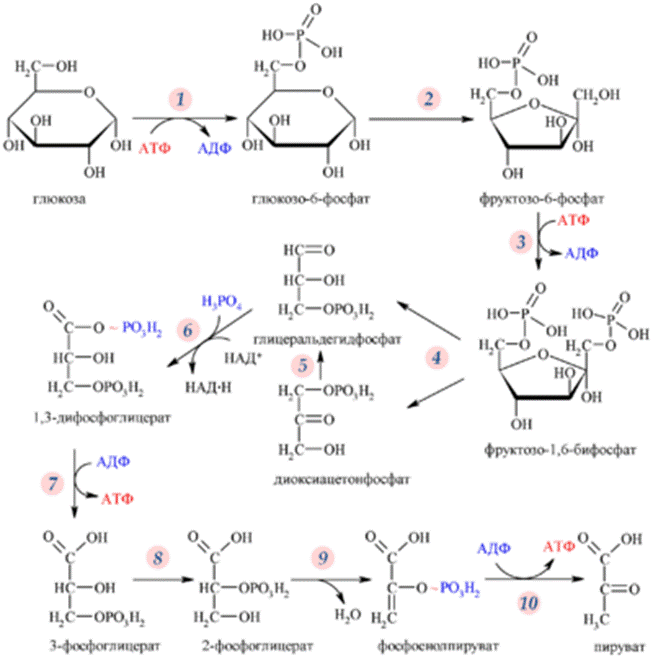
Вторая стадия гликолиза включает реакции фосфорилирования, в ходе которых свободная энергия, содержащаяся в исходной молекуле глюкозы, высвобождается и запасается в форме АТР.
6 реакция. Эта стадия начинается с окисления глицеральдегид-3-фосфата с образованием высокоэнергетического соединения 1,3-бифосфоглицерата путём присоединения фосфата. Эту окислительно-восстановительную реакцию катализирует глицеральдегид-3-фосфатдегидрогеназа.
В результате этой сложной реакции альдегидная группа глицеральдегид-3-фосфата окисляется, но при этом образуется не карбоновая кислота, как можно было ожидать, а смешанный ангидрид фосфорной и 3-фосфоглицериновой кислот - 1,3-бифосфоглицерат. Такого типа ангидрид, называемый ацилфосфатом, характеризуется очень высоким значением стандартной свободной энергии гидролиза ацилфосфатной связи -49,3 кДж/моль.
7 реакция. В следующей реакции 1,3-бифосфоглицерат используется для образования АТР; фосфатная группа высокоэнергетического ацилфосфата с участием фосфоглицераткиназы переносится на АДР с образованием АТР и 3-фосфоглицерата.
AG0 реакции переноса = - 49,3 + (+30,5) = - 18,8 кДж/моль, иными словами эта реакция сильно сдвинута в сторону образования АТР.
В ходе последующих реакций гликолиза происходит образование пирувата и второй молекулы АТР.
8 реакция. Фосфоглицератмутаза катализирует перенос фосфатной группы от кислорода при С-3 на кислород при С-2, в результате чего 3-фосфоглицерат превращается в 2-фосфоглицерат.
9 реакция. Далее осуществляется реакция дегидрирования 2-фосфоглицерата до фосфоэнолпирувата. Реакция катализируется енолазой (фосфопируватгидратазой).
Несмотря на сравнительно небольшое изменение стандартной свободной энергии в ходе данной реакции, величина AG0 гидролиза фосфатной группы исходного вещества и продукта различается очень сильно. Для 2-фосфоглицерата - низкоэнергетического фосфорилированного соединения, эта величина равна 17,5 кДж/моль, а для фосфоэнолпирувата - высокоэнергетического фосфорилированного соединения - 61,9 кДж/моль.
10 реакция. Образованный фосфоэнолпируват отдаёт фосфатную группу АДР при участии пируваткиназы. Оставшийся при этом енол самопроизвольно превращается в пируват.
AG0 этой реакции = -61,9 + (+30,5) = - 31,4 кДж/моль.
Глюкозо-6-фосфат может образовываться и без затраты АТР путём отщепления гликозильных остатков с помощью специфических фосфорилаз. В частности у животных в печени и в скелетных мышцах находится фермент гликогенфосфорилаза, который катализирует фосфоролиз 1,4-гликозидной связи нередуцирующих концов гликогена; глюкозные остатки отщепляются один за другим в форме глюкозо-1-фосфата. Глюкозо-1-фофат, образующийся из гликогена при участии фосфоглюкомутазы превращается в глюкозо-6-фосфат, дальнейшая судьба которого в печени и в мышцах различна. В печени глюкозо-6-фосфат превращается в глюкозу при участии глюкозо-6-фосфатазы, глюкоза уходит в кровь и используется в других органах и тканях. В мышцах нет этого фермента и глюкозо-6-фосфат распадается аэробным или анаэробным путём.
11 реакция. Восстановление пирувата до лактата. При аэробных условиях в животных тканях продуктом гликолиза является пируват, НАДН, образовавшийся в ходе окисления глицеральдегид-3-фосфата, реокисляетя (т.е. снова превращается в НАД+ за счёт молекулярного кислорода. В аэробных условиях, например в напряжённо работающих скелетных мышцах, НАДН реокисляется за счёт пирувата, который при этом восстанавливается в лактат. Это означает, что НАД может использоваться в гликолизе многократно. Эта реакция катализируется ферментом лактатдегидрогеназой.

Полный баланс гликолиза. В анаэробных условиях одна молекула глюкозы превращается в две молекулы лактата (путь углерода). Две молекулы АДР и две молекулы фосфата превращаются в две молекулы АТР (путь фосфатных групп). Четыре электрона в форме двух гидрид-ионов переносятся с помощью двух молекул НАД+ от двух молекул глицеральдегид-3-фосфата на две молекулы пирувата с образованием двух молекул лактата (путь электронов).

Две молекулы NADH в аэробных условиях вновь окисляются до NAD+, дыхательной цепью митохондрий. Здесь электроны передаются в конечном счёте на кислород.
Регуляция гликолиза. Скорости главных катаболических реакций, обеспечивающих расщепление глюкозы и извлечение химической энергии в форме АТР регулируются в соответствии с потребностями клетки в АТР независимо от использования АТР. Поскольку продукты расщепления глюкозы играют важную роль и в качестве предшественников и как промежуточные продукты других метаболических процессов, регуляторные ферменты катаболизма углеводов распознают также соответствующие сигналы других метаболических путей и отвечают на эти сигналы.
Вовлечение глюкозных остатков в процесс гликолиза обеспечивают две важные реакции, и обе эти реакции контролируются регуляторными ферментами. Первая реакция - катализируется гексокиназой. В некоторых тканях, например в скелетных мышцах, гексокиназа функционирует как аллостерический фермент и ингибируется продуктом реакции глюкозо-6-фосфатом.
Во второй реакции, поставляющей глюкозные остатки для процесса гликолиза, субстратом служит гликоген. Эта реакция катализируется гликоген-фосфорилазой, которая также представляет собой регуляторный фермент. В скелетных мышцах этот фермент присутствует в двух формах - в каталитически активной фосфорилированной форме (фосфорилиза а) и в значительно менее активной дефосфорилированной форме (фосфорилиза b). Взаимопревращения этих двух форм гликоген-фосфорилазы происходит под действием специфичных ферментов, катализирующих процесс ковалентной модификации фосфорилизы - фосфатазы (ингибируется ионами кальция и AMP) и киназы (активируется ионами кальция).
Особая роль в выполнении регуляторной функции гликолиза отводится фосфофруктокиназе. Превращение фруктозо-6-фосфата в фруктозо-1,6-бифосфат, катализируемое этим ферментом, является наиболее медленно текущей реакцией гликолиза, определяющей скорость всего процесса. Фосфофруктокиназа, будучи аллостерическим ферментом, активируется фосфатом, AMP, АДР, ингибуруется АТР и цитратом.
Пируваткиназа активируется фосфоенолпируватом и АДФ; активируется цитратом и АТР.
Следует однако иметь ввиду, что у разных организмов регуляторные этапы гликолиза могут значительно отличатся характером регуляции.
Не нашли, что искали? Воспользуйтесь поиском: