ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
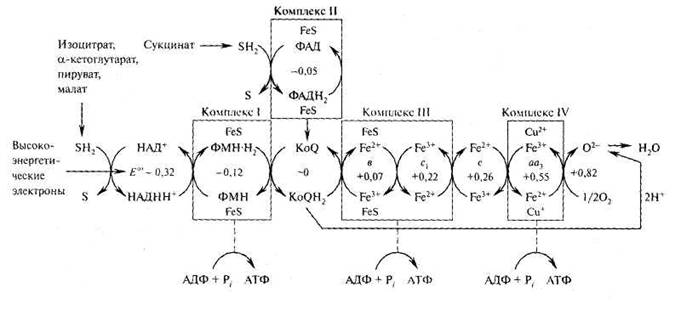
Транспорт электронов и окислительное фосфорилирование.
Все ферментативные этапы окислительного расщепления углеводов, жиров и аминокислот сходятся в аэробных клетках к конечной стадии клеточного дыхания – окислительному фосфорилированию, на которой электроны переходят от органических субстратов к кислороду, а энергия, выделяемая при этом, используется для образования АТР из ADP и фосфата.
В каждом обороте ЦТК специфичные дегидрогеназы отщепляют от изоцитрата, α-кетоглутарата, сукцината и малата четыре пары атомов водорода. Эти атомы водорода в определенной точке отдают свои электроны в цепь переноса электронов и превращаются таким образом в ионы Н+, которые поступают в водную среду. Кроме того, образуются и другие атомы водорода, отщепляемые дегидрогеназами от пирувата, жирных килот и аминокилот во время расщепления этих соединений до ацетил-СоА и других продуктов. Они все передают свои электроны в дыхательную цепь к конечному акцептору электронов – кислороду.
Дыхательная цепь состоит из ряда белков с прочно присоединенными простетическими группами, обладающими способностью присоединять и отдавать электроны. Эти белки располагаются в определенной последовательности, в которой каждый из них способен присоединять электроны от предыдущего и передавать их тому, который следует за ним.
Компоненты дыхательной цепи расположены во внутренней мембране митохондрий. Внутреннее пространство митохондрии окружено двумя мембранами (рис.). Наружная мембрана гладкая, а внутренняя образует многочисленные складки или кристы. Пространство, ограниченное внутренней мембраной, заполнено матриксом, который примерно на 50% состоит из белка. У прокариот дыхательная цепь локализована в структурах цитоплазматической мембраны.
Переносчики электронов, ферменты и некоторые мембранные транспортные системы, находящиеся во внутренней мембране митохондрий, составляют до 75% от общей массы мембраны.
В настоящее время экспериментально определена последовательность расположения переносчиков электронов в дыхательной цепи (рис.). Окислительно-восстановительный потенциал переносчиков электронов в этой последовательности постепенно становится все более положительным.
В 60-х гг. XX в., благодаря методам мягкого разрушения интактных митохондрий, были выделены четыре дыхательных комплекса (I, II, III, IV):
комплекс I (НАДН: KoQ-оксидоредуктаза) катализирует перенос электронов от НАДН к KoQ;
комплекс II (сукцинат: KoQ-оксидоредуктаза) - перенос электронов от сукцината к KoQ;
комплекс III (KoQH2: цитохром с-оксидоредуктаза) - перенос электронов от KoQH2 к цитохрому с;
комплекс IV (цитохромоксидаза) катализирует перенос электронов от цитохрома с к кислороду.
Выделение комплексов дыхательной цепи позволило сделать вывод об определенной пространственной ориентации этих комплексов в мембране. Важная роль в передаче электронов от одного комплекса к другому принадлежит KoQ и цитохрому с. Цитохром с является единственным растворимым цитохромом и наряду с коэнзимом Q служит мобильным компонентом дыхательной цепи, осуществляя связь между фиксированными в мембране комплексами.
Реакции переноса электронов – это окислительно-восстановительные реакции. Соединения, отдающие электроны в такой реакции, называются донорами электронов, или восстановителями, а соединения, присоединяющие электроны, - акцепторами, или окислителями.
Окислители и восстановители всегда функционируют как сопряженные окислительно-восстановительные пары (редокс-пары). Способность каждой сопряженной окислительно-восстановительной пары обратимо отдавать электрон выражают стандартным окислительно-восстановительным потенциалом Е0 (или просто стандартный восстановительный потенциал). Чем более отрицательной величиной выражается восстановительный потенциал системы, тем выше ее способность отдавать электроны; и наоборот, чем более положительной величиной выражается восстановительный потенциал, тем выше способность системы присоединять электроны.
Процесс переноса электронов связан с синтезом АТР. Сопряжение реакций окисления восстановленных молекул (НАДН • Н+ или ФАДН2) и образования АТР и называется окислительным фосфорилированием.
Впервые представление о сопряжении между аэробным дыханием и фосфорилированием АДФ было высказано в 30-х гг. XX столетия В.А. Энгельгардтом.
Для количественной оценки сопряжения окисления и фосфорилирования используют коэффициент сопряженного окислительного фосфорилирования (Р/О), предложенный в 1939 г. В.А. Белицером. Коэффициент Р/О — это отношение уменьшения числа молей неорганического фосфата (Н3РО4), необходимого для синтеза АТФ, к количеству поглощенного кислорода. Так, для субстратов (малат, пируват, изоцитрат), окисляющихся НАД-зависимыми дегидрогеназами, Р/О = 3, а для сукцината, окисляемого ФАД-сукцинатдегидрогеназой, Р/О = 2.
Процесс дыхания не всегда является процессом, жестко сопряженным с фосфорилированием. Нарушают систему сопряжения процессов окисления в дыхательной цепи и фосфорилирования так называемые разобщающие агенты (разобщители). К ним относятся вещества, подавляющие синтез АТФ (фосфорилирование), в то время как окисление субстратов, потребление кислорода (дыхание) продолжаются. В качестве разобщителей в экспериментальной биохимии используют 2,4-динитрофенол, динитрокрезол, пентахлорфенол и др. В присутствии разобщителей коэффициент Р/О равен нулю, а энергия окисления в этом случае трансформируется в тепловую форму. Следовательно, разобщители обладают пирогенным действием, т.е. повышают температуру тела. Большинство разобщающих агентов являются липофильными и их ингибирующее действие на процесс фосфорилирования легко объяснимо благодаря способности этих соединений обеспечить протонную проводимость сопрягающей мембраны митохондрий и тем самым препятствовать образованию электрохимического потенциала, а следовательно, и синтезу АТФ.
Факторами, лимитирующими скорость дыхания, являются доступность кислорода, АДФ, субстратов, возможности и состояние самой дыхательной цепи при насыщающих концентрациях всех субстратов и компонентов.
Не нашли, что искали? Воспользуйтесь поиском: