ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Вит. В6 и РР. Роль в обмене АК, примеры р-ций, строение.

Участвует в образовании НАД и НАДФ.
1. Компонент дыхательной цепи.
2. КОФЕРМЕНТ различных дегидрогеназ.
Суточная потребность в витамине РР 20 - 25мг.
Источниками витамина РР являются дрожжи, говяжья печень, рыба, грибы, мука пшеничная, соя, бобы, хлеб, картофель, мясо. Может синтезироваться в организме при поступлении с пищей белков и витамина В6. Гиповитаминоз проявляется в виде пеллагры:
1. Дерматит с повреждением симметричных участков кожи, повреждённых УФО.
2. Диарея.
3. Деменция.
Причины гиповитаминоза: белковое голодание, недостаток витамина В6.

Участвует в образовании ПФ:
1. КОФЕРМЕНТАМИНОТРАНСФЕРАЗ.
2. КОФЕРМЕНТДЕКАРБОКСИЛАЗ.
3. Принимает участие в ДЕЗАМИНИРОВАНИИ.
4. Необходим для образования витамина РР из триптофана.
5. Сигма-АМИНОЛЕВУЛИНОВАЯ кислота ® гем.
Т.о. витамин В6 участвует в обмене аминокислот, следовательно, необходим для нормального
обмена белков. Суточная потребность-2мг. Источники вит.В6: картофель, пшеница, рис, отруби, печень, дрожжи. Гиповитаминоз приводит к нарушению белкового обмена, что проявляется развитием анемии, дерматита, стоматита, глоссита.
100. Хар-ка вит. С, строение. Участие в обмене в-в, проявление гиповитаминоза. Вит. Р. Витамин С (аскорбиновая к-та, антискорбутный).
Химическая структура:

L-аскорбиновая кислота. L-дегидроаскорбиновая кислота
Витамин С легко окисляется в нейтральной и щелочной среде в присутствии кислорода. Метаболические функции витамина С.
1.Участие в окислительно-восстановительных реакциях:
антиоксидант, гидроксилирование катехоламинов,
ДОФА ® НОРАДРЕНАЛИН
гидроксилирование аминокислот, входящих в состав коллагена
ПРОЛИН ® ОКСИПРОЛИН
2.Синтез кортикостероидов
Суточная потребность в витамине С - 100 - 120мг.
Источниками витамина С являются: лимон, грецкие орехи, смородина, шиповник, яблоки, красный перец, картофель, зелёный лук. укроп, квашеная капуста. Дефицит вит.С приводит к активации свободно радикальных процессов (стресс, ОРВИ). Гиповитаминоз проявляется в виде нарушения синтеза коллагена: увеличивается проницаемость сосудов, следовательно, кровоточивость десен, патехии на коже, кариес. Причины гиповитаминоза:
1. высокая потребность,
2. неустойчивость химической структуры,
3. отсутствие депо.
Витамин Р (витамин проницаемости, РУТИН, КАТЕХИНЫ, ПОЛИФЕНОЛЫ).
Обладает антиоксидантной активностью, участвует в окислительно-восстановительных реакциях.
Тормозит активность гиалуронидазы - фермента, разрушающего гиалуроновую кислоту
(компонент соединительной ткани). Суточная потребность не установлена. Источниками витамина Р являются апельсины, лимон, шиповник, смородина, грецкий орех, салат, томат, капуста, картофель, чай, сухое красное вино. Гиповитаминоз проявляется аналогично гиповитаминозу С.
101. Вит. В12 и фолиевая к-та. Их хим. природа, участие в метаболических процессах. Причины гиповитаминозов. Витамин В12 (кобаломин, антианемический).
По структуре геминоподобное соединение. Устойчив при нагревании, но не образуется ни в организме человека, ни в организме животных, ни в растениях, но синтезируется микроорганизмами.
Метаболические функции витамина В12.
В процессе метаболизма из КОБАЛОМИНА образуются МЕТИЛКОБАЛОМИН и АДЕНОЗИНКОБАЛОМИН. МЕТИЛКОБАЛОМИН участвует в транспорте метильных групп
I.МЕТИЛИРОВАНИЕ В12
ГОМОЦИСТЕИН ® МЕТИОНИН ® ХОЛИН ® ФОСФОТИДИЛХОЛИН
2.ДЕМЕТЕЛИРОВАНИЕ В12
МЕТИЛТЕТРОГИДРОФОЛЕВАЯ К-ТА ® ТЕТРОГИДРОФОЛЕВАЯ К-ТА
II.АДЕНОЗИЛКОБАЛАМИН В12
МЕТИЛМАЛОНИЛ-КОА ® СУКЦИНИЛ-КОА
Суточная потребность в витамине 1,5-Змкг. В клинике используют до 200мкг. В этом случае он усваивается без внутреннего фактора КАСТЛА. Источниками витамина В12 являются печень говяжья и трески, мясо, сыр, яйцо, молоко коровье, козье. Витамин В12 синтезируется микрофлорой кишечника. Он депонируется в печени. Его запасов хватает на 2 - 3 месяца.
Гиповитаминоз В12:
1.Клетки теряют фолевую кислоту, а её дефицит проявляется в виде мегалобластической анемии.
2.Поражение слизистой ЖКТ.
3.Накопление МЕТИЛМАЛОНИЛ-КОА. следовательно, повышение синтеза жирных кислот с
нечётным количеством атомов углерода.
4.Нарушение синтеза ХОЛИНА и ФОСФОЛИПИДОВ, следовательно, повышается синтез нейтральных жиров, приводящее к жировой инфильтрации печени.
5.Повышение ГОМОЦИСТЕИНА в крови приводит к развитию атеросклероза.
Причины гиповитаминоза. Для всасывания витамина В12 необходим МУКОПРОТЕИН (фактор КАСЛА), который синтезируется обкладочными клетками слизистой желудка.
1. Недостаточность или нарушение рецепторов.
2. Дефицит мукопротеина (иммунные расстройства, заболевания слизистой желудка).
3. Дефицит витамина В12 (длительное вегетарианство, энтериты, дсбактериозы).
В крови витамин В12 транспортируется с помощью белка ТРАНСКОБАЛОМИНА. Выделяют его несколько форм. С помощью транскобаломина-2 витамин транспортируется в печень и там депонируется, связывается с транскобаломином-1 и поступает снова в кровь. Транскобаломин-3 предупреждает потери витамина. Курение понижает активность витамина В12.
102. Витамины – антиоксиданты, их био роль. Редоксвитамнны или витамины - антиоксиданты - это вещества, которые препятствуют развитию процессов свободно-радикального окисления. Это природные оксиданты: Е, С, А.
103. Биотин, пантотеновая к-та, их роль в обмене в-в. Витамин Н (биотин, антисеборейный).
Метаболические функции витамина Н
1.Является КО-ферментом карбоксилаз ПВК, ацетил -КОА, пропионил-КОА.
ПВК + CО2 (вит.Н) ® ЩУК
2.Участвует в реакциях синтеза жирных кислот и стерина.
Суточная потребность в витамине Н 0,15 - 0,2мг. Источниками витамина Н являются печень, соя, молоко, яйца, мука, лук, морковь, апельсины, дрожжи, арахис. Синтезируется микрофлорой кишечника. Гиповитаминоз проявляется в виде чешуйчатого дерматита (носогубной треугольник и волосистая часть головы), конъюктевита, анемии, себореи. Причины гиповитаминоза: дисбактериозы. Употребление сырых яиц. содержащих АВИДИН, связывающий витамин Н, и этот комплекс не усваивается. Заболевания ПЖЖ, в которой синтезируется фермент БИОТИНИДАЗА, освобождающий биотин от белка; если этого фермента нет, то БИОТИН не усваивается.
ПАНТОТЕИНОВАЯ КИСЛОТа (витамин ВЗ или В5).
Является производной бета -АЛАНИНА, соединенной с производным масляной кислоты. Метаболические функции ПАНТОТЕИНОВОЙ кислоты.
1.Входит в состав КО-фермента А, следовательно, участвует в синтезе АЦЕТИЛ-КОА, различных АЦИЛ-КОА, образующихся в результате следующих реакций:
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ альфа –КЕТОКИСЛОТ. Синтез и окисление жирных кислот, синтез СТЕРОИДОВ.
2.Участвует в синтезе более 80 различных ферментов.
Суточная потребность 10-15мг. Источники: печень, дрожжи, пчелиное молочко. Синтезируется микрофлорой кишечника. Гиповитаминоз характеризуется поражением -малых -артерий нижних конечностей. В эксперименте наблюдался некроз надпочечников.
104. Витаминоподобные в-ва. Антивитамины. ВИТАМИНОПОДОБНЫЕ ВЕЩЕСТВА.
Синтезируются в организме, но в недостаточном количестве. Потребность в них измеряется в граммах. Например, витамин F, содержащий три незаменимые аминокислоты (ЛИНОЛЕВУЮ, ЛИНОЛЕНОВУЮ, АРАХИДОНОВУЮ). Его относят в настоящее время к витаминподобным веществам. Т.к. суточная потребность в нём до 10гр.
ПАБК (ПАРААМИНОБЕНЗОЙНАЯ К-ТА)

1. Участвует в образовании ФОЛИЕВОИ кислоты,
2. Участвует в образовании ряда ферментов,
3. Является фактором пигментации.
Недостаточность ПАБК проявляется в виде нарушения пигментации. Суточная потребность не установлена. Источники: печень, дрожжи и другие продукты.

1. Участвует в образовании ФОСФОТИДИЛХОЛИНА.
2. Донор - СНЗ групп для образования ПУРИНОВЫХ и ПИРИМИДИНОВЫХ оснований.
3. Необходим для образования АЦЕТИЛ ХОЛИ НА.
Суточная потребность: 0,5 - 1 гр. Источники: желток яиц, печень, почки и др. продукты.
ОРОТОВАЯ КИСЛОТА (является пиримидиновым основанием)

Необходима для синтеза ПИРИМИДИНОВЫХ И ПУРИНОВЫХ азотистых оснований, предшественников аминокислот. Содержится в дрожжах и печени.
ПАНГАМОВАЯ КИСЛОТА - производное глюкуроновой кислоты и диметилглицина. В организме быстро гидролизуется. ИНОЗИТ (производное циклогексана)

Входит в состав ФОСФОТИДИЛИНОЗИТА. При его недостатке развивается дерматит и жировое перерождение печени. Суточная потребность 1 -2гр.
АНТИВИТАМИНЫ - это вещества, нарушающие усвоение витаминов или понижающие биологическую активность витаминов.
По действию различают АНТИВИТАМИНЫ:
1.Прямо воздействующие:
белок яйца АВЕДИН + БИОТИН не усваиваются ТИАМИНАЗА - разрушение тиамина.
2.Структуры аналогичные витаминам:

СА включается в ферменты микроорганизмов, не отличающие их по структуре. Функции ферментов нарушается, и микроорганизмы погибают.
МЕТОТРИКСАН - антивитамин фолиевой кислоты. Используется как противоопухолевый препарат, снижает белок синтетические процессы в клетках.
ДИКУМАРИН - антивитамин К, снижающий свёртываемость крови.
ФТИВАЗИД, ТУБАЗИД - антивитамин В6.
105. Сигнальные молекулы и хим. частицы, их классификация. Виды регуляторных эффектов сигнальных молекул. Факторы роста. Отличительные признаки гормонов. Классификация гормонов. Понятие о клетке мишени. Роль гипоталамуса в гормональной регуляции. Виды регуляции обмена в-в. Внешняя регуляция. Основные задачи регуляции метаболизма и клеточных функций:
1.внутриклеточное и межклеточное согласование клеточных процессов,
2.исключение «холостых» циклов метаболизма, продукты которых не востребованы,
3.эффективное образование и использование энергии,
4.поддержание гомеостаза,
5.приспособление организма к условиям окружающей среды.
Регуляция метаболизма: внутренняя и внешняя. Внутренняя регуляция - управляющие сигналы образуются и действуют внутри одной и той же клетки (само-регуляция). Внешняя регуляция - управляющие сигналы поступают к клетке из внешней среды. Внутренняя регуляция осуществляется путём изменения активности ферментов активаторами или ингибиторами. Особенно активно при внутренней регуляции работают аллостерические ферменты (ФОСФОРИЛАЗА). Внешняя регуляция обеспечивается специализированными сигнальными молекулами - эндогенные химические соединения, которые в результате взаимодействия с ферментами, обеспечивают внешнее управление биохимическими процессами в клетках-мишенях.
Клетка-мишень - это клетка, имеющая специализированные воспринимающие рецепторы для данного вида сигнальных молекул. Сигнальные молекулы являются лигандами для рецепторов клеток-мишеней.
Характерные особенности сигнальных молекул.
1.малый период жизни (динамичность, оперативность регуляции).
2.высокая биологическая активность (действие развивается при очень низких концентрациях).
3.уникальность, неповторимость действия. Эффекты одного типа сигнальных молекул не могут быть смоделированы другим. Это обеспечивает разнообразие регуляции.
4.наличие эффекта усиления (одна сигнальная молекула может усиливать каскады биохимических р-ий).
5.один вид сигнальных молекул может иметь несколько клеток-мишеней.
6.реакция разных клеток-мишеней на одну и ту же сигнальную молекулу отличается (объясняется многообразием рецепторов и их своеобразием).
Способы внешнего управления клетками-мишенями.
1.Управление экспрессией генов (биосинтез белков и ферментов). Это медленный способ регуляции.
2.Управление активностью ранее синтезированных белков:
- управление активностью ферментов, следовательно, изменение биохимических процессов;
- изменение активности функциональных белков, следовательно, прямое изменение функции клеток. Например, влияние сигнальных молекул на ионный канал вызывает деполяризацию мембраны и формирование потенциала действия. Чрезвычайно быстрый эффект.
Виды регуляторных эффектов сигнальных молекул:
1.Эндокринный. Сигнальные молекулы поступают с током крови из желудочно-воротной системы к клеткам-мишеням. Так действует большинство гормонов.
2.Паракринный - сигнальные молекулы вырабатывают в пределах одного органа или участка ткани. Таким образом действуют большинство факторов роста.
3.Аутокринное - сигнальные молекулы действуют на клетку, их образовавшую.
КЛАССИФИКАЦИЯ СИГНАЛЬНЫХ МОЛЕКУЛ.
1)По химической природе:
1.Органические (производные аминокислот, жиров). СТЕРОИДЫ, ПРОСТОГЛАНДИНЫ.
2.Неорганические - 1992г. МОНООКСИДАЗОТА (NO).
2)По физико-химическим свойствам:
1.Липофобные - не могут проникать через мембрану клетки. Они растворимы в воде.
2.Липофильные - растворяются в жирах. Свободно проникают через ЦПМ и действуют на рецепторы внутри клетки. Например, производные холестерина: МИНЕРАЛО-, ГЛЮКОКОРТИКОИДЫ, ЭСТРОГЕНЫ, АНДРОГЕНЫ, ЙОДТИРОНИНЫ, NO.
3)По биологическому принципу:
1.Гормоны - сигнальные молекулы с выраженным эндокринным эффектом.
2.Цитокины - факторы роста. Это сигнальные молекулы белковой природы, которые выделяются неспециализированными клетками организма. Они регулируют рост, дифференцировку, пролиферацию соседних клеток. Действие пара- и аутокринно.
3.Нейромедиаторы сигнальные молекулы, вырабатывающиеся нервными клетками, координирующие работу нейронов и управление периферическими тканями. Их действие связано с влиянием на ионные каналы. Они изменяют их проницаемость и вызывают деполяризацию мембраны.
106. Вторые посредники в действии липофобных сигнальных молекул, цАМФ и цГМФ – зависимые механизмы действия. Аденилатциклаза, протеинкиназа. Продемонстрировать эффекты гормонов, осуществляющие регуляторное действие при участии цАМФ. Механизм действия ЛИПОФОБНЫХ сигнальных молекул:
1. взаимодействие с поверхностными рецепторами,
2. сигнал передаётся от рецептора внутрь клетки (ТРАНСДУКЦИЯ) и устанавливается с помощью внутриклеточных регуляторов: высоко- и низкомолекулярных.
Высокомолекулярные регуляторы - это регуляторные белки. Они опосредуют действие сигнальной молекулы внутри клетки.
Низкомолекулярные регуляторы небелковой природы. Его называют второй МЕССЕНДЖЕР (первый МЕССЕНДЖЕР - сама сигнальная молекула) - полноправный представитель сигнальной молекулы внутри клетки. Это ионы кальция, ДИАЦИЛГЛИЦЕРОЛ, ИНОЗИТОЛТРИФОСФАТ, цАМФ и цГМФ.
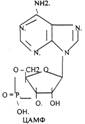
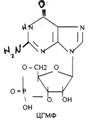
3. биологическое действие обусловлено сочетанием регуляции активности ранее синтезированных белков и регуляция экспрессии генов. Регуляторный эффект двухфазный:
- первая фаза быстрая, но не продолжительная, она обеспечивает изменение структуры и активности ферментов;
- вторая фаза медленная за счёт изменения количества ферментов.
МЕХАНИЗМ ДЕЙСТВИЯ, ЗАВИСИМЫЙ ОТ ЦАМФ.
Факторы, необходимые для этого:
1. растворимая в воде сигнальная молекула;
2. поверхностные рецепторы клетки-мишени;
3. внутриклеточный трансдуктор G-белок. Состоит из 3 единиц: альфа, бета, гамма.
G-белок может быть ингибирующий и активирующий. Он находится в непосредственной близости к рецептору. При взаимодействии сигнальной молекулы с рецептором он активируется, его активность обусловлена альфа единицей. Она оказывает влияние на внутриклеточный фермент - АДЕНИЛАТЦИКЛАЗУ (превращает АТФ в ЦАМФ). Альфа-S повышает активность. альфа-I понижает активность G-белка. G-белок способен присоединять ГДФ или ГТФ. Альфа-единица активна, когда связана с ГТФ.
4. АДЕНИЛАТЦИКЛАЗА(АЦ);
5. ПРОТЕИНКИНАЗА-А ЦАМФ-зависимая. Она катализирует реакцию фосфорилирования белков. В результате белки изменяют активность;
6. Регуляторные элементы ДНК (ЭЕХАНСЕР и САЙЛЕНСЕР);
7. ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ;
8. ФОСФАТАЗА - дефосфорилируют белки;
9. Белок-синтетический аппарат клетки.
Этапы, стимулирующие ЦАМФ -зависимый механизм:
1. взаимодействие сигнальной молекулы с рецептором;
2. изменение конформации G-белка;
3. замена ГДФ на ГТФ в альфа-S единице G-белка;
4. альфа-S ГТФ активирует АЦ;
5. АЦ синтезирует ЦАМФ;
6. ЦАМФ активирует ПРОТЕИНКИНАЗУ-А (ПКА);
7. ПКА фосфорилирует белки и белковые факторы транскрипции, изменяющие активность и количество ферментов;
8. Прекращение действия, если рецептор освободился - альфа-5-С-ГТФ-азная активность:
альфа-единица может разрушать ГТФ - это таймер действия.
- ФОСФОДИЭСТЕРАЗА - разрушает ЦАМФ.
- ФОСФАТАЗА - ДЕФОСФОРИЛИРУЕТ белки. Этапы, ингибируюшие ЦАМФ -зависимый механизм:
С первого по третий те же самые этапы, отличие в G-белке (альфа-I единица). Четвёртый этап - связывание ГТФ с альфа-I единицей будет ингибировать АЦ. Ингибируюший механизм противодействует и прекращает эффекты ЦАМФ в клетке.
ЦГМФ -зависимый стимулирующий механизм действия.
Рецептор встроен в мембрану клетки и связан с ферментом ГУАНИЛАТЦИКЛАЗОЙ (ГЦ). При присоединении сигнальной молекулы ГЦ активируется и катализирует реакцию ГТФ * ЦГМФ. Последний активирует ПРОТЕИНКИНАЗУ-G (ПКО), а она запускает реакцию фосфорилирования белков (ферментов и факторов транскрипции).
107. Механизм действия липофильных сигнальных молекул. Механизм действия NO. Принципы иммунноферментного анализа уровня сигнальных молекул. Механизм действия ОРГАНИЧЕСКИХ ЛИПОФИЛЬНЫХ сигнальных молекул.
1.взаимодействие с внутриклеточными рецепторами,
2.регуляторный эффект связан с изменением количества белков в результате влияния на экспрессию генов (действуют через геном),
3. биологическое действие продолжительное, но развивается медленно в пределах часов.
Факторы, необходимые для их действия:
- сигнальные молекулы,
- воспринимающий внутриклеточный рецептор, связанный с шапероном.
- участок ДНК, регулирующий транскрипцию определённых генов (ЭНХАНСЕР, САЙЛЕНСЕР),
- белок синтетический аппарат клетки.
Этапы действия:
1. проникновение внутрь клетки,
2. связывание с внутриклеточным рецептором,
3. освобождение шаперона (запуск таймера действия),
4. взаимодействие комплекса сигнальных молекул с регуляторными элементами ДНК, изменение биосинтеза некоторых белков, в том числе и их ферментов.
5. изменение метаболизма и клеточных функций.
Механизм прекращения действия органических липофильных сигнальных молекул:
1. разрушение рецепторов, обусловленное отсутствие защиты со стороны шаперона,
2. протеолиз синтезированных белков,
3. разрушение факторов транскрипции, участвующих в передаче сигналов к структурному гену.
По перечисленному механизму действуют СТЕРОИДНЫЕ гормоны и ЙОДТИРОНИН.
Механизм действия НЕОРГАНИЧЕСКИХ ЛИПОФОБНЫХ сигнальных молекул (NO). NO беспрепятственно проникает через мембрану клетки. Это короткоживущая молекула, образующаяся в организме под действием NO-СИНТАЗЫ из аминокислоты АРГ. В клетке NO взаимодействует с ГЦ, активирует её, что вызывает накопление в клетке ЦГМФ. который активирует ПКО, и развивается клеточный ответ по выше рассмотренному механизму.
Эффекты NO:
1. фактор расширения сосудов;
2. регулятор АПОПТОЗА (запрограммированной клеточной смерти);
3.NO является свободным радикалом, поэтому способен влиять на процесс перекисного окисления жиров и регулировать функции МИТОХОНДРИЙ;
4. является ИММУНОМОДУЛЯТОРОМ.

Не нашли, что искали? Воспользуйтесь поиском: