ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Карбонатное дыхание
Карбонатное дыхание ведут ацетогенные и метаногенные бактерии. Они в качестве акцептора электронов используют СО2 и восстанавливают его: метаногенные до метана, ацетогенные до ацетата.
Серное дыхание
Некоторые бактерии способны расти в присутствии элементарной серы, используя ее в качестве конечного акцептораэлектронов. При этом сера восстанавливается до сероводорода. Пример: Desulfuromonasacetoxidans.
19 вопрос. Фотосинтезирующие микроорганизмы. Пигменты. Световая и темновая фазы фотосинтеза. Механизм фиксации углекислоты автотрофными фотосинтезирующими прокариотами. Фотофосфорилирование. Две фотосинтетиченских системы в зависимости от экзогенных доноров электронов. Циклический и нециклический перенос электронов при фотофосфорилировании.
фотосинтези́рующие бакте́рии-осуществляют фотосинтез при участии пигментов – хлорофиллов, используя энергию света. У одних фотосинтез сопровождается выделением кислорода (оксигенные бактерии), у других нет (аноксигенные фототрофы). Оксигенные бактерии представлены цианобактериями, они, как все водоросли, содержат хлорофилл а. Аноксигенные фототрофы содержат различные формы бактериохлорофиллов, к ним относят зелёные и пурпурные серные бактерии, пурпурные несерные бактерии.
Зелёные и пурпурные серные бактерии в фотосинтезе вместо воды используют сероводород, пурпурные несерные бактерии в качестве донора электронов используют органические соединения. Пурпурные серные бактерии внутри своих клеток запасают серу, встречаются в пресных и солёных водоёмах, серных источниках, образуя пурпурные налёты на иле или окрашенные слои воды. Среди них есть как подвижные, так и неподвижные формы. Зелёные серные бактерии – группа очень мелких бактерий разной морфологии. Живут в строгих анаэробных условиях, серу в клетках не накапливают, а выделяют её наружу. Большинство способно к азотофиксации. Большое количество пигментов с широкой цветовой гаммой продуцируют грибы, в т. ч. патогенные для человека. П., растворимые в воде, окрашивают колонии и среду; не растворимые в воде П. окрашивают только колонии. П. фотосинтезирующих бактерий наряду с бактериохлорофиллом принимают участие в дыхании. У гетеротрофных организмов они выполняют функцию защиты от повреждающего действия видимых и УФЛ.
СВЕТОВÁЯ ФÁЗА ФОТОСИ́НТЕЗА [. Часть процессов фотосинтеза, непосредственно зависящая от энергии света. С.ф.ф. начинается с поглощения кванта света молекулой одного из светособирающих пигментов. Далее энергия возбужденной светом молекулы передается на хлорофилл реакционного центра, где преобразуется в энергию движущихся электронов. Сопряжение транспорта электронов с переносом ионов водорода внутрь тилакоида создает предпосылки для последующего синтеза АТФ. Именно фотосинтетическое фосфорилирование, осуществляемое на мембранах тилакоидов хлоропластов является центральной частью С.ф.ф. Во время С.ф.ф. с необходимостью происходят и др. важнейшие события, напр. фотолиз воды. Основными же продуктами этого этапа являются НАДФН и АТФ. С.ф.ф. противопоставляется темновая фаза фотосинтеза.ТЕМНОВÁЯ ФÁЗА ФОТОСИ́НТЕЗА. Ферментативные реакции, связанные с усвоением углерода углекислого газа растением. Т.ф.ф. не нуждается непосредственно в световой энергии, если в хлоропластах имеются в достаточном кол-ве продукты световой фазы фотосинтеза АТФ и НАДФН. Ядром Т.ф.ф. является цикл Кальвина. Т.ф.ф. противопоставляется световая фаза фотосинтеза.
Фотоавтотрофные (фотосинтезирующие) эубактерии. В качестве источника углерода используют углекислый газ. Для восстановления углекислого газа используют световую энергию. В клетках содержатся фотосинтетические пигменты: бактериохлорофиллы. Донорами электронов для восстановления углекислого газа служат органические вещества, водород, сероводород. Анаэробы: акцепторами электронов при дыхании служат органические окислители, при этом полного окисления органических веществ не происходит. Представители: фотосинтезирующие пурпурные серные бактерии, зеленые бактерии. В то же время есть данные, что у зеленых бактерий, а также у некоторых пурпурных бактерий (R. rubrum) может функционировать особый путь ассимиляции углекислоты, названный восстановительным циклом карбоновых кислот или циклом Арнона. В этом цикле углекислота фиксируется на органических кислотах, причем две реакции, а именно карбоксилирование ацетил-КоА и сукцинил-КоА, происходят при участии восстановленного ферредоксина. В условиях интенсивного роста фототрофных бактерий большая часть первичных продуктов ассимиляции углекислоты и органических соединений используется на синтез белков и других важнейших компонентов клеток. Но при задержке роста культур в результате недостатка фосфора и особенно азота происходит образование в больших количествах запасных веществ. И пурпурные и зеленые бактерии могут накапливать в качестве запасных продуктов полисахариды типа гликогена. Фотофосфорилирование -Одна из форм фосфорилирования АМФ и АДФ с образованием АТФ, свойственная фотосинтезирующим организмам; Фотофосфорилирование происходит в процессе фотосинтеза с использованием энергии видимого света.
Фотохимическая стадия фотосинтеза обеспечивает поднятие электрона на высокий энергетический уровень и начало транспорта его через ряд переносчиков с постепенно снижающимися окислительно-восстановительными потенциалами. Перенос электрона может происходить по циклическому и нециклическому пути. В первом случае он возвращается к исходному донору, т. е. к пигменту фотоактивного центра; при втором этого не происходит. Поэтому для поддержания нециклического транспорта электронов необходим экзогенный Н-донор, который восполняет их потерю. Обе такие системы функционируют при фотосинтезе у растений. Перенос электрона по циклическому пути, как и в дыхательной цепочке, сопряжен с синтезом АТФ. Механизм этих процессов, видимо, одинаков. В результате транспорта электронов по нециклическому пути, кроме АТФ, происходит образование восстановителей, исцользуемых при ассимиляции углекислоты и других соединений. Эту функцию выполняют восстановленный ферредоксин И (НАДН и НАДФН). АТФ и указанные восстановители рассматриваются как первые стабильные продукты фотосинтеза, которые иногда называют «ассимиляционной силой». При бактериальном фотосинтезе также имеет место циклический путь переноса электронов, сопряженный с образованием АТФ. Но способ образования восстановителей не вполне ясен. Не исключено, что их образование, как и у растений, происходит в результате функционирования нециклической фотосинтетической системы транспорта электронов. Природа Н2А может быть разной. У растений такую функцию выполняет вода. Поэтому фотосинтез сопровождается выделением кислорода. У бактерий Н-донором служат сероводород, сера, тиосульфат, молекулярный водород или органические соединения. Поэтому кислород не выделяется, а накапливаются продукты их окисления. Возвращение электрона - темновой процесс. Электрон перемещается по цепи переносчиков в соответствии с электрохимическим градиентом. Имеет место циклический транспорт электронов.
Циклическим электронным транспортом у фотосинтезирующих эубактерий не исчерпываются все возможные пути переноса электронов. Электрон, оторванный от первичного донора реакционного центра, может по цепи, состоящей из других переносчиков, не возвращаться к молекуле хлорофилла, а передаваться на такие клеточные метаболиты, как НАД(Ф)+ или окисленный ферредоксин, которые используются в реакциях, требующих восстановителя.
Таким образом, электрон, покинувший молекулу хлорофилла, выводится из "системы". Возникает однонаправленный незамкнутый электронный поток, получивший название нециклического пути переноса электронов. У пурпурных и зеленых нитчатых бактерий функционирует только циклический светозависимый поток электронов. У остальных групп эубактерий фотоиндуцируется как циклический, так и нециклический перенос электронов, при этом у зеленых серобактерий и гелиобактерий оба пути электронного транспорта связаны с функционированием одной фотосистемы, а у цианобактерий и прохлорофит циклический перенос электронов зависит от активности фотосистемы I, a для нециклического потока электронов необходимо функционирование обеих фотосистем. Поток электронов по цепи переносчиков на определенных этапах сопряжен с направленным перемещением протонов через мембрану, что приводит к созданию протонного градиента, используемого для синтеза АТФ.
Вопрос. Брожение. Сбраживаемые и несбраживаемые субстраты.Получение энергии путем субстратного фосфорилирования. Реакции и ферменты субстратного фосфорилирования.Основные типы брожения. Окислительная и восстановительная фазы брожения.
Брожение- это ферментитивный метаболический процесс превращения органических соединений в отсутствии кислорода, в котором синтезируется АТФ, а продукты расщепления органического субстрата служат одновременно донорами и акцепторами электронов. Исходя из определения, а какие же это органические соединения.
При брожениях конечными акцепторами электронов служат в основном органические соединения: метаболиты, образующиеся из исходных субстратов(пировиноградная кислота, ацетальдегид), или вещества, имеющиеся в среде культивирования (некоторые аминокислоты и другие органические соединения, способные восстанавливаться). В ряде брожений акцепторами электронов служат молекулы СО2, а также ионы водорода(Н+). Брожение-это способ получения энергии, при котором АТФ образуется в процессе анаэробного окисления органических субстратов в реакциях субстратного фосфорилирования.
1. Возможность частичного окисления и восстановления субстрата в результате внутримолекулярного расщепления.
2. Субстрат превращается в продукт с выделением энергии.
3. Микроорганизмы, живущие в энергии брожения они всегда 2-х стадийные:
1 стадия: окислительное
2 стадия: кислорода нет; восстановительная стадия протекает без образования АТФ.
4. Пути расщепления сахаров.
4.1. Гликолиз у факультативных облигатных анаэробов.
4.2. Пентозофосфатный у многих прокариот, которые факультативно анаэробы.
4.3. КДФГ: встречается редко, но в основном у аэробов. Есть исключения.
Как получается энергия?
5. Субстратное фосфорилирование =фосфатная группа + АДФ. Фосфат отщепляется от субстрата.
5.1. 1,3- фосфоглицераткиназа
5.2. Фосфоенолпируваткиназа
5.3. Ацетилфосфаткиназа
5.4. Бутирилфосфаткиназа
Окисление идёт дальше, образование 4-х углеродного соединения, АДФ идёт дальше.
6. Какие субстраты могут быть сбражены.
Сбраживаемые субстраты: полисахариды, гексозы, пентозы, тетрозы, многоатомные спирты, органические кислоты, аминокислоты(исключения: ароматические кислоты, которые сбраживаются частично, где есть О и Н), азотистые соединения: пурины и пиримидины.
7. Несбраживаемые субстраты: длинные цепочки углеводородов, алифатические и ароматические углеводороды, стероиды, каратиноиды, терпены и порфирины.В аэробных условиях все эти вещества поддаются расщеплению и полностью окисляются, но в анаэробных условиях они очень стабильны. Стабильность их может быть обусловлена двумя причинами. 1. Большинство названных соединений содержит только атомы углерода и водорода; при внутримолекулярном расщеплении таких веществ энергия не выделяется. 2. Насыщенные углеводороды и полиизопреноиды могуть окисляться только в присутствии молекулярного кислорода; первичное воздействие на них катализируется в этом случае оксигеназой.
Как эти вещества разлагаются и как можно извлечь энергию?
Могут подвергаться аэробному окислению (под действием особых ферментов, ферменты, включения О2 в несбраживаемых соединениях.
Включения- монооксигиназы-О
-диоксигиназы-О2 (в молекулу несбраживаемого субстрата)
8. Основные типы брожения:
Спиртовое брожение: пировиноградная кислота в результате её окислительного декарбоксилирования образуется ацетальдегид, который становится конечным акцептором водорода. В итоге из 1 молекулы гексозы образуются 2 молекулы этилового спирта и молекулы углекислоты. Процесс получил название спиртового брожения.
Процесс спиртового брожения суммарно можно выразить следующим образом:
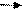 С6Н12О6+ 2Фн+2АДФ 2СН3-СН2ОН+2СО2+2АТФ+2Н2О.
С6Н12О6+ 2Фн+2АДФ 2СН3-СН2ОН+2СО2+2АТФ+2Н2О.
1 молекула глюкозы приводит к образованию 2 молекул АТФ.
Происходит межмолекулярное размежевание на восттановленные (этиловый спирт) и окисленные (СО2) молекулы.
Дрожжи: S.cerevisiae, Kloeckra. Sarcina venriculi, Zymomonas mobilis.
Молочное брожение:
Гомоферментативное молочнокислое брожение. Гомоферментативные молочнокислые бактерии образуют практически только одну молочную кислоту. Катаболизм глюкозы происходит у них по фруктозобисфосфатному пути, а водород, отщепляющийся при дегидрирование глицеральдегид-3-фосфата, передается на пируват. Лишь небольшая часть пирувата декарбоксилируется, превращаясь в уксусную кислоту, этанол и СО2, а также в ацетоин. Количество образующихся побочных продуктов зависит, по-видимому, от доступа кислорода.
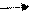 С6Н12О6 2СН3-СНОН-СООН
С6Н12О6 2СН3-СНОН-СООН
Streptococcus lactis, S. faecalis, L.lactis,S. cremoris, Pediococcus cerevisiae и т.д.
Гетероферментативное молочнокислое брожение. При гетероферментативном молочнокислом брожении образуется больше продуктов: молочная кислота, уксусная кислота, этанол, двуокись углерода. Тип брожения, в основе которого лежит гексозомонофосфатный путь сбраживания пентоз или гексоз с образованием в качестве акцепторов электронов пирувата и ацетилфосфата. Пируват восстанавливается до лактата (СН3-СНОН-СООН),а ацетилфосфат, в зависимости от активности энергетического метаболизма бактерий, у одних (род Lactobacillus) превращается а ацетат(СН3СООН) и СО2, а у других (род Leuconostoc) – через ацетил-КоА и ацетальдегид (СН3-СНО) восстанавливается в этанол(С2Н2ОН). Энергетический выход процесса при образовании этанола составляет одну молекулу АТФ на одну молекулу окисленной глюкозы, а при образовании ацетата-две молекулы АТФ.
Leuconostoc mesenteroides, Lactobacillus brevis и т.д.
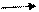 Брожение маслянокислое- тип брожения, осуществляемый «сахаролитическими» анаэробными клостридиями-Clostridium butyricum,C. Pasteurianum и другими, сбраживающими крахмал, целлюлозу и пектин по пути гликолиза с образованием пирувата (СН3-СО-СООН), который подвергается ферредоксинзависимому окислительному декарбоксилированию с образованием ацетил-КоА.Далее в реакции конденсации типа С2+С2 С4 из двух молекул ацетил-КоА формируется четырехуглеродный скелет-ацетоацетил-КоА. Дальнейшие превращения ацетоацетил-КоА сводятся к реакциям дегидратации, восстановления, декарбоксилирования и дезактивации. В результате этих реакций доминирующими продуктами брожения становятся бутират (СН3-СН2-СН2-СООН), ацетат (СН3-СООН), СО2 и Н2. Энергетический выход маслянокислого сбраживания глюкозы составляет три молекулы АТФ(две образуется на стадии гликолиза и одна на стадии регенерации ацетата).
Брожение маслянокислое- тип брожения, осуществляемый «сахаролитическими» анаэробными клостридиями-Clostridium butyricum,C. Pasteurianum и другими, сбраживающими крахмал, целлюлозу и пектин по пути гликолиза с образованием пирувата (СН3-СО-СООН), который подвергается ферредоксинзависимому окислительному декарбоксилированию с образованием ацетил-КоА.Далее в реакции конденсации типа С2+С2 С4 из двух молекул ацетил-КоА формируется четырехуглеродный скелет-ацетоацетил-КоА. Дальнейшие превращения ацетоацетил-КоА сводятся к реакциям дегидратации, восстановления, декарбоксилирования и дезактивации. В результате этих реакций доминирующими продуктами брожения становятся бутират (СН3-СН2-СН2-СООН), ацетат (СН3-СООН), СО2 и Н2. Энергетический выход маслянокислого сбраживания глюкозы составляет три молекулы АТФ(две образуется на стадии гликолиза и одна на стадии регенерации ацетата).
Уксуснокислое брожение. Уксуснокислым брожением называется окисление этилового спирта в уксусную кислоту под влиянием уксуснокислых бактерий. Оно может быть выражено таким суммарным уравнением:
С2Н5ОН +О2= СН3СООН+ Н2О
Это брожение, как и спиртовое, известно с давних времен. Человек с давних пор наблюдал, что на поверхности вина или пива, оставленных в открытом сосуде, образуется сероватая пленка, а содержимое превращается в уксус. Микробиологическая природа этого процесса была впервые установлена в 1862 г. Пастером.
Возбудителями уксуснокислого брожения являются уксуснокислые бактерии, составляющие многочисленную группу палочковидных, бесспоровых, аэробных бактерий. Среди них встречаются подвижные и неподвижные формы. Различаются они также размерами клеток, разной устойчивостью к спирту и способностью накапливать больше или меньше уксусной кислоты.
Ruminococcus albus и т.д.
Основные продукты пропионовокислого брожения, вызываемого несколькими видами бактерий из рода Propionibacterium, — пропионовая (CH3CH2OH) и уксусная кислоты и CO2. Химизм пропионовокислого брожения сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например, в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём брожения. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной кислоты и CO2 образуется щавелевоуксусная кислота, превращающаяся в янтарную кислоту, из которой декарбоксилированием образуется пропионовая кислота.
Основное энергетическое значение для пропионовокислых бактерий имеют так называемые ключевые реакции пропионовокислого брожения.
Под пропионовокислым брожением подразумевают биохимический процесс превращения бактериями сахара, молочную кислоту и ее солей в пропионовую кислоту. В этом брожении, кроме пропионовой кислоты, образуются и такие продукты, как уксусная кислота, углекислый газ, янтарная кислота, ацетоин, диацетил, другие летучие ароматические соединения - диметилсульфид, ацетальдегид, пропионовый альдегид, этанол и пропанол. Химизм данного брожения подобен типичному молочнокислому брожению с той разницей, что образовавшаяся молочная кислота в этом брожении не конечный продукт, а промежуточный. От других типов брожения пропионовокислое отличается высоким выходом АТФ, участием некоторых уникальных ферментов и реакций.
Propionibacterium freudenreichii
Муравьинокислое брожение, или смешанное брожение-тип брожения, вызываемый энтеробактериями и протекающий по пути гликолиза с образованием газов(Н2,СО2), кислот(ацетат, лактат, сукцинат, формиат) и нейтральных (ацетоин, этанол, 2,3 –бутандиол) продуктов, главным из которых выступает формиа(НСООН).1. Пируват подвергается расщеплению с образованием формиата и ацетил-КоА, который превращается в ацетат. Это позволяет энтеробактериям ассимилировать энергию в форме АТФ путем фосфорилирования АДФ на уровне субстрата. Энергетический выход процесса составляет три молекулы АТФ. 2. Ацетолактат превращается в ацетоин, который может восстановиться до 2,3-бутандиола, либо окисляться в диацетил. Этанол образуется из ацетил-КоА, а СО2 и Н2-из формиата.
К энтеробактериям относятся прдеставители филы Proteobacteria: Proteus, Serratia, Escherichia, Klebsiella.
Не нашли, что искали? Воспользуйтесь поиском: