ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Клеткалық иммунитет жүйесі. Иммунологиялық төзімділік. Цитокиндер.
60-жылдарда Дж.Миллердин тышқандарға тимоэктомия тәжірибе жасаумен
перифериялық мүшелерде Т-лимфоциттердің саны азайғаны және аллогенді трансплантаттың ұзақ отырып қалуы, антиденелердің өндіруінің бұзылуы табылған.
Сүйек кемігінің пре-Т-лимфоциттері тимустың субкапсулярлық қабатында бөлініп мүшеленеді. Функция жөнінде бұл клеткалардың ішінде СД4, СД8 маркерлардың тасымалдаушылары айырылады. Эпителиалды клеткалармен өндірілген биологиялық заттардың көмегімен Т-лимфоциттер даму процесін өтіп қыртысты қабатқа миграциялайды. Қыртысты қабатта жағымды селекциядан өтеді, яғни І-ІІ классты гистосәйкестік антигендерге сәйкес рецепторлары бар клондарды сұраптайды. І классты МНС танушы клондар – Т-киллерлер болып түзіледі, І, ІІ классты МНС танушы клондар Т-хелперлер болады.
Т-клеткалардың сыртында тек қана бір корецептор (СД3) экспрессорлайды. Ол антиген байланыстыру рецепторлардың антигендермен өзара әрекетін күшейтеді және сигналды клетка ішіне жеткізу ролін атқарады. СД4 және СД8 позитивтік клеткалар Т-хелпер мен Тс ретінде саналады. Аутоантигендермен әсерлесу қабілеті бар клеткалар апоптоз жолымен жойылады. Сұраптау шарттан өткен клеткалар иммун жүйесінің перифериясына барады.
Т-клеткалардың В-жүйеден айырмасы – Т-клеткалар антигенді жеке танымайды, яғни І-ші немесе ІІ-ші МНС-ның молекулаларымен комплекс құрап отырып таниды. І-ші классты молекуласы кіретін комплексті цитотоксикалық СД8 Т-клеткалар таниды, ал ІІ-ші классты молекуласы кіретін комплекс СД4 Тн1 қабыну клеткаларымен немесе СД4 Тн2 хелперлы клеткалармен танылады.
Эффекторлы клеткалардың генерациясы.
Антигенді біріншілік тануы аңқау Т-клеткалармен жүзеге асады. Аңқау Т-клетканың жетілуі лимфоидты тканьде өтеді. Лимфа ағымымен патоген жақын лимфа түйініне, ал қан ағымымен көк бауырға барады. Антиген кілегей қабықтарға орналасқанда пейер табақшаларға, бадамша бездерге миграциялайды.
Перифериялық лимфоидты тінде АГ танып жэне оны Т-лимфоциттерге иммуногенді түрде ұсынуға қаблеті бар 3 типті мамандандыру клеткалар бар. Олар – макрофагтар, дендритті клеткалар және В-клеткалар. Бұл клеткалардың функциясы 1-ші немесі 2-ші МНС молекулалардың антигенді пептидтермен комплексті түзуімен көрінеді. Антиген лимфоидты тінге түсіп лимфоциттердін рециркуляциясын күшейтеді. Аңқау Т-лимфоциттер лимфотүйіндердің Т-аймағына венуланың эндотелийі арқылы түседі. Жетілген Т-клеткалардың генерациясы макрофагтардың және дендритті клеткалардың бетінде пептидті антигенді І немесе ІІ-ші МНС классты молекулалармен комплексте танудан басталады. Кофакторлардың әсері Т-клеткалар рецепторларының антигендік комплекспен әсерлесуіне мүмкіндік береді. Осы қызметті АТК-лар жүзеге асырады.
Аңқау Т-клеткалардың антигенмен біріншілік кездесуден кейін активациясы “сәйкестендіру” (примирлеу) деп аталады. Осыдан кейін Т-клеткалардың әрбір субпопуляцияларының қызметтік бағыттары анықталады. Егер аңқау СД4 ТнО клеткалар антигендік комплексті макрофагтың бетінде таныса, ары қарай СД4 Тн1 клеткаларға мүшеленеді. Егер МНС ІІ-ші молекуламен комплексте антиген В-клетканың бетінде танылса, ары қарай СД4 Тн2 хелперлі клеткаларға мүшеленеді.
Аңқау Т-клеткалардың көпшілігі арнайылық әсерлесудің сұрыптауынан өте алмайды, қайтадан рециркуляция процесіне қатысу үшін, өзіне тән антигенді іздеп эфферентті лимфа тамырлары арқылы кетеді.
Т-клеткалардың антигентанушы рецепторларының антигендік комплекспен байланысуы және СД8 немесе СД4 корецепторлардың қатысуы, аңқау Т-клеткалардың дамуына бірінші сигналды қамтамасыз етеді – пролиферациямен дифференцировкаға. Даму процесінің жалғасуына екінші сигнал қажет. Костимулятор ретінде АПК мембранасында экспрессерленетін В7 молекуласы шығады. В7 – гомодимер, иммуноглобулиндердің супер тұқымдасына жатады. Аңқау Т-клеткаларадың бетінде В7 рецепторы ретінде СД28 белок болып табылады. В7: СД28 өзара әсерлесуі екінші сигналды қамтамасыз етеді.
Активтелген аңқау Т-клеткалар СД28-ден басқа костимулдеуші активтілігі бар тағы бір белокты синтездеп экспрессерлейді – CTLA-4. Ол СД28-ге қарағанда В7-ге аффинділігі жоғары.
СД4 Тн1-ші қабыну клеткалардың активтілігі:
Тн1-ші қабыну клеткаларының қатысуымен макрофагтардың активтенуіне 2 сигнал қажет: 1-ші – СД4 Тн1-ші қабыну клеткаларымен түзілетін ИНФ-γ; 2-ші – макрофагтарды белсендіру үшін беткей ФНО- α қызмет етеді. Нәтижесінде макрофагтарда биохимиялық өзгерістер жүреді, осы клеткалар жоғары антибактериалды қасиет көрсетеді. Фагоцитоз процесі оттектік жарылыспен қоса жүреді – оттектік радикалдар, азот тотығы түзіліп бактерицидті белсенділігін көрсетеді. Сонымен қатар активтелген макрофагтар МНС І және ІІ классты молекулаларының, ФНО- α рецепторының экспрессиясын күшейтеді. Бұл процесс қосымша аңқау клеткалардың араласуына әкеледі. Макрофагтармен әсерелесетін қабыну Т-клеткалар өздері де белсеніп, иммунды жауаптың ұйымдастырушы ролінде шығады.
НК-клеткалар
Вируспен инфицирленген клеткаларды жою реакциясына қатысады. Бұл ірі лимфоциттер Т- және В-клеткалардың маркерларысыз, антиген танушы рецепторлары жоқ, антигенмен әсерлесуден кейін пролиферацияға, иммунологиялық есте сақтау қабілеті жоқ. Бірақ, Т-клеткалар цитокиндердің әсерімен активтенеді. НК-клетка бетінде IgG1 және IgG2 Ғс-фрагментіне арналған рецептордың болуы, олардың осы антиденелерге сәйкес антигендермен әсерлесуін болжайды. Бұл антидене тәуелді клеткалық байланысты цитотоксикалық әсер.
Т-жүйесінің қызметтері.
1. Клетка ішінде орналасқан патогендерден қорғау, трансплатациялық, ісікке қарсы иммунитетке, созылмалы қабынуға қатынасу. Клеткалық иммунитет цитотоксикалық Т-лимфоциттермен, активтелген макрофагтармен орындалады. Микробтар клетка ішінде орналасқан жұқпалы ауруларда гранулема пайда болады. Ол макрофаг, Т-лимфоциттер, эозинофилдер жиынынан құралады. Гранулема микробты ошағынан шығармайтын жергілікті созылмалы қабыну процессі.
Клеткалық иммунитеттің негізгі реакциялары:
1. СД8 клеткалардың цитотоксикалы киллерлік әсері.
2. Баяу сезімталдықтың дамуы.
Вирусты инфекцияда, ісік өскен кезде, трансплантатты қабылдамау
реакциясында басты эффекторлар ретінде цитотоксикалық СД8 Т-клеткалар болады.
Т-киллердің нысана клеткалармен әрекеттік әсерінің кезеңдері: 1) І МНС
молекуламен біріккен антиген комплексті бастаушы клеткалармен тануы, олардың пролиферациясы және дифференциясы арқылы эффекторларға дейін жетілуі; 2) леталды соққы беру; 3) лимфотоксин (ФНО-β) көмегімен клетканы бүлдіру және киллердің ағытылуы. СД8 клеткалардың сонымен қатар супрессиялық қабілеті болады.
Баяу сезімталдық – антиген қайталанып, енген орнында 24-28 сағаттан кейін дамитын қабыну реакциясының жоғары сезімталдық түрі.
Антиген интактты организмге бірінші рет түскенде макрофагпен жұтылады, клетка ішінде ыдырайды және макрофагтың бетінде МНС ІІ класс молекуласымен комплексте экспрессерленеді. Бұл иммуногенді комплекс аңқау СД4 Т-клеткалармен танылады (Тн0). Олар ары қарай СД4 Тн1 клеткаларға мүшеленеді. Иммунды жауапқа қатынасқан Т-лимфоциттер екіншілік иммунды жауапты қамтамасыз етілу үшін еске сақтау клеткаларын түзейді.
Антиген қайталап енген кезде, жетілген Тн1 қабыну клеткалар антиген тану реакциясына түседі. Танудың нәтижесінде Тн1 клеткалар антиген енген аймаққа макрофагтарды және қабынудың басқа клеткаларын жақындататын хемотаксикалық факторларды бөледі.
Клиникада клеткалық иммунитеттке баға беру:
1. Шеткері қанда Т-лимфоциттердің және олардың субпопуляцияларының санын моноклоналды антидене және розетка түзу эдістер көмегімен анықтау. Моноклоналды антиденелермен анқтау мүмкіншіліг жоқ жерде Т-лимфоциттердің жалпы санын қой эритроциттерімен розетка түзу арқылы анықтайды.
2.Т-лимфоциттердің функционалды қызметін анықтау үшін тесттер:
-митогенге және арнайы антигенге Т-лимфоциттердің бласттрансформация реакциясы – РБТЛ
- Т-лимфоциттердің белгілі субпопуляцияларымен синтезделетін лимфокиндердің өнімін мөлшерін өлшеу “ МИФ, ИӘ-2, 5,6,13, ИНФ-± және т.б.).
Иммунологиялық төзімділік – арнайы антигенге иммунды жауапты тудыру қабілетін жоғалту.
Төзімділіктің дамуы – мүшелерді қондырғанда өте қажет. Бірақта, қатты жұқпалы ауруларды қиын салдарға әкеледі.
Табиғи және жасанды төзімділіктің механизмдері бірдей.
Иммунологиялық тозімділікті қамтамасыз ету факторлары:
-Иммунитеттін орталық мүшелерінде (тимуста) клондардың теріс сұрыпталу нәтижесінде.
--АТК клеткалардын қызметінің төмендеуі. Олар В7 белогын синтездемейді.
- Аутоантигендермен әсерлесу қабілеті бар Т- және В-клеткаларды жоюлуы,
- Т- және В-лимфоциттердің корецепторларын блоктау,
- Аутореактивті клондардың жұмысын басатын супрессор клеткалар қызметінің төмендеуы.
-Антиидиотипті антиденелердің қатысуы.
Иммунологиялық төзімділік ерігіш антигендерге, эмбрионалды және неонаталды кезеңдерде, антигеннің оралды жолдармен, көп немесе өте төмен мөлшерінде енгізгенде, АГ-нің құрылысы қарапайым болса, әлсіз АГ-ге қарсы, жетілмеген Т-және В-лимфоциттермен әрекеттескенде женіл пайда болады. Антиген антитоксинмен байланысқанда арнайы Т-мен В-клеткаларды зақымдайжы. АГ мен АД комплекстер рецепторларды блоктап, ісікке қарсы клеткалық иммунитеттің реакциясын басуы мүмкін, гаптендерді иммуногендік емес тасымалдаушымен блоктауы төзімділікке әкеледі.
Цитокиндер.
Клеткалық және гуморальді жауап түзетін, клетка аралық механизмдер мен иммунды компонентті клеткалардың дифференцировкасын зерттеген кезде, белокты медиаторларының үлкен тобы ашылды, олар цитокиндер деп аталады. Олардың 30-дан астам түрлері белгілі.
Цистокиндердің негізгі биологиялық белсенділігі- иммунды жауапты барлық даму сатыларында реттеу: функциональді белсенді иммунокомпонентті клеткалардың пролиферациясы мен дифференцировкасы, антигенді өңдеу және макрофагтардың бетіңде иммуногенді түрде ұсыну, антиген сезімтал лимфоциттердің пролиферациясы, иммуноглобулиндер продуценттеріне дейін В- клеткалардың дифференцировкасы, бір изотоптан басқасына иммуноглобулиндер синтезін ауыстыру, цитотоксикалық Т- клеткалардың ізашаларының эффекторларға дейін пісіп жетілуін қамтамасыз ету, макрофагтардың цитотоксикалық әсерін индукциялау. Кейбір цитокиндер қабыну реакциясына және ісік клеткаларының элиминациясында айқындала түсетін жедел фазалы жауапқа белсенді түрде қатысады.
Құрылыс ерекшеліктері мен биологиялық әсеріне қарай барлық цитокиндер бірнеше өзіндік топтарға бөлінеді: гемопоэтиндер, интерферондар, иммуноглобулиндердің супер топтық цитокиндері, ФНО- тобы цитокиндері, хемокиндер.
Цитокиндер өзара 5 топқа бөлінеді.
1-ші топ – Гемопоэтикалық цитокиндер.
1. ГМ-КСФ – гранулоцитарлы моноцитарлы клеткалардың ізашарларын стимулдейді.
2. М-КСФ – моноциттердің ізашарларын стимулдейді.
3. Г-КСФ – гранулоциттердің ізашарларын стимулдейді.
4. ИЛ-3 – мульти КСФ
5. ИЛ-5 – эозинофилдерге арналған КСФ
6. ИЛ-7 – лимфоциттерге арналған КСФ
7. ИЛ-1 – ең ерте қан түзуші клеткалардың ізашарларын стимулдейді.
2-ші топ – Иммундыққа дейінгі қабынудың цитокиндері:
а) біріншілік про-қабыну цитокиндер – ИЛ-1, ТНФ-α, ИЛ-6. Макрофагтармен және дендритті клеткалармен бөлінеді. ИЛ-1-ші мен ТНФ-α жергілікті әсер етеді, ал ИЛ-6 жетілік фаза белоктардың синтезін бауырда стимулдейді.
Б) екіншілік қабыну цитокиндер – ол лейкоциттер мен лимфоциттердің локоматорлы жылжуын қамтамасыз етеді.
3-ші топ – Лимфоцитарлы иммунды жауаптың ұйымдастырушылары.
Т- және В-, НК-клеткаларының пролиферациясымен дифференцировкасын реттейді. Бұл цитокиндерді антиген АТК клеткалар және лимфоциттер бөледі. Оларға жататындар: ИЛ-2, ИЛ-4, ИЛ-12, ИЛ-15 және ИФН-γ жатады.
4-ші топ – Иммунды қабынудың медиаторлары. Т-лимфоциттермен бөлінеді, лейкоциттерді қабынуға активтейді: ИФН-γ (макрофагтармен НК-клеткалардың активаторы), ИЛ-5 (эозинофилдердің индукторымен активаторы), LT (нейтрофилдердің активаторы), LT-α (қабынулық гранулалардың пайда болуын қамтамасыз етеді).
5-ші топ – Қабынуға қарсы цитокиндер.
ИЛ-10 – макрофагтармен бөлінеді және оларды тежейді.
TGF-β – иммунды СД4 Т-лимфоциттермен бөлінеді және лимфоциттердің ары қарай пролиферациясын тежейді. Сонымен қатар ИЛ-4 пен ИЛ-13 макрофагтармен ингибирлеуші әсер етеді.
Интерлейкин- 1
ИЛ-1 өнімінің негізгі көзі түрлі тіндік локализациясының фагоциттеуші мононукулеаларлар болып табылады: перифериялық қан мен перитонеальді экссудаттың макрофагтары мен моноциттері, бауырдың купфер клеткалары, эпидермистегі Лангерганс клеткалары, нервті тіннің микроглия клеткалары.
ИЛ-1- дің белсенді туындыларына эндотелиоциттер жатады. Одан басқа, бұл цитокинді секреттеу қабілетіне Т және В- лимфоциттер, фибробласттар, НК- клеткалар, кератиноциттер, нейтрофильдер ие.
Тыныштық күйдегі макрофагтар, және цитокиннің басқа клеткалық көздері, ИЛ-1-ді бөлмейді және оның мРНК-сы жоқ.
ИЛ-1-геннің экспрессиясы биологиялық белсенді белоктың түзілуімен жүретін, түрлі индукторлар арқылы клеткаларының активациясынан кейін ғана басталады. ИЛ-1-дің продукциясын шақырушы заттар арасында бактериялардың клеткалық қабырғасының компонентері мен қорғаныс реакциясының дамуы барысында пайда болатын цитокиндер болып келеді.
ИЛ-1-дің маңызды ең бір қасиеті Т- лимфоциттердің пролиферациясын ынталандыру болып табылады. ИЛ-1-дің өз бетімен Т-лимфоцидтердің өсуі үшін фактор бола алмайды. Оның әсер ету механизмі, Т- хелперлермен секреттелетін, ИЛ-2 және ИЛ-4 өсу факторларының, синтезін күшейту.
Сонымен қатар, Ил-1 Т- хелперлердің пролифеорациясын аутокринді түрде реттеу үшін жағдай туғызып, ИЛ-2 және ИЛ-4 рецепторларының экспрессиясын күшейтеді. ИЛ-1-ң ынталандырушы белсенділігі ИЛ-4 өңдейтін Т- хлеперлермен байлансты (Тн2). Иммунды жауаптың дамуы үшін ИЛ-1 дің В- клеткаларға өсуін- стимульдеуші әсерінің маңызы зор.
Арнайы антигендермен немесе митогиндермен белсендірілген В- клеткалар берілген цитокиннің әсерінен пролиферациясын күшейтеді. Бірақ, Т- хелперлер сияқта, бұл стимульдеуші әсері ИЛ-2-ге арналған рецепторларының экспрессиясын белсендіру арқылы әсер етеді. Сонымен қатар В- лимфоциттер дифференцировкасында ИЛ-1-дің қатысуы маңызды. Ол өздігінен дифференцирлеуші белсенділік қасиетіне ие емес, бірақ басқа цитокиндермен бірлесіп, антиденепродуциентті клеткаларға примирленген клеткалардың трансформациясын қамтамасыз етеді.
ИЛ-1 арнайы иммундық жауапқа қатысудан басқа, қорғаныстың арнайы емес формаларының дамуына жауапты- инфекциялық зақымдану кезіндегі жедел фазалы жауап және жергілікті қабыну реакциясының түзілуі деңгейінде ең маңызды медиаторлар ретінде қатысады.
Интерлейкин-2
Ең бірінші анықталған цитокин- интерлейкин-2(ИЛ-2). Ил-2-нің негізгі продуциенттері Т- хелперлер болып табылады. Бұл типті клеткалардың 75% Ил-2 синтездейді. Цитотоксикалық Т- клеткалардың 20%-на дейін бұл цитокинді бөлуге қабілетті. Бұл клеткалардағы ИЛ-2 синтезіне антигендер немесе митогендер ғана әсер етпейді, басқа да биологиялық белсенді қосылыстардың да әсер бар. Басқа клеткалар кластарымен бөлінетін белгілі цитокиндер (ИЛ-1,ИЛ-6,ФНО,ИФН), антигендермен белсендірілге Т-клеткалармен ИЛ-2-нің продукциясын ынталандырады. Тимус гармондары (тимозин, тимустың сары сулық факторы) ИЛ-2 продуцент клеткаларына жетілмеген тимоциттердің дифференцировкасын қамтамасыз етеді. Клетка ішілік Са2+ деңгейін күшейтуші ионофорлар жаңағы цитокиннің продукциясын күшейтеді.
ИЛ-2- нің реттеуші әсерінің нысанасы болып Т- клеткалардың субпопуляциялары, В- клеткалар, табиғи киллерлі клеткалар, макрофагтар болып табылады.
ИЛ-2 әсерінің негізі нәтижесі тыныштық күйдегі немесе антиген және митоген арқылы ынталандырылған клеткалардың пролиферациясын қамтамасыз етеді. Осы ИЛ-2-нің биологиялық белсенділігі лимфо- миелоидты комплексті клеткалардың өспелі фактор ретінде анықтайды.
Интерлейкин- 3
Интерлейкин-3(ИЛ-3)- қан түзуші клеткалардың ерте ізашарлары үшін өспелі және дифференцирлеуші фактор.
ИЛ-3-тің негізгі продуценті Тhy-1 және Тhy-2, сонымен қатар цитотоксикалық Т- лимфоциттердің бір бөлігі мен басқа клеткалар (тимустың эпителиальді клеткалары, В- лимфоцитер, миелоидтты және сүйек кемігінің страмалды клеткалары). Жаңағы цитокин барлық гемопоэтикалық өспелі дифференцировкалардың ерте ізашарларынан түрлі фенотипті клеткалардың жетілуіне мүмкіндік жасайды: лимфоидты, моноцитарлы, гранулоцитарлы, мегакариоцитарлы, эритроидты. ИЛ-3-тің Т- лимфоциттердің ерте ізшараларына дифференцирлеуші әсер етуін ескеру қажет. Осындай әсер ету нәтижесінде жетілмеген Тhy-1 Тhy-1+ Л- лимфоциттерге ауысады. Бұл цитокин пре- В- клеткалардың беткей иммуноглобулинді экспрессиясының басталуына дейінгі дифференцировкасына қатысады.
Интерлейкин-4
Ил-4-тің көзі митогенмен стимульденген Т- хелперлер (Тhy-2), мес клеткалар, сүйек кемігінің стромасының индентифицирленбеген клеткалары болып табылады. Сәйкес рецепторлары бар ИЛ-4-тің реттеуші әсерінің нысанасына Т және В лимфоциттер, макрофагтар, фибробласттар, НК- клеткалар, мес клеткалар, гемопоэздің сүйек кемігінің ізшаралары жатады. ИЛ-4 В-клеткалардың пролиферациясына костимулятор ретінде қатысады. Сонымен қатар ИЛ-4 Jg Е- нің продукциясының деңгейін жоғарлатуға қабілетті. ИЛ-4 макрофактормен синтезделетін қабынуды шақыратын цитокиндердің (IL-1 b, 6, 8, 12, TNF –a) түзілуін және азот пен от тегінің метоболиттерінің пайда болуын тежейді. ИЛ-4 лимфокин арқылы активтелген клеткалардың (ЛАК) және макрофагтардың ісікке қарсы активтілігін күшейтеді. ИЛ-4 секрециясының реттелуінің бұзылуы аллергопатологияның дамуында маңызды рол атқарады. Созылмалы С-вирусты гепатитінде ИЛ-4-тің жоғарлауы анықталады, әсірісе асқыну кезеңінде.
Интерлейкин-5
Бұл цитокиннің негізгі продуценттері митоген немесе антигенмен стимулденген Т- хлеперлер (Тhy-2) болып табылады. Тыныштық күйдегі клеткалар ИЛ-5 үшін генді экспрессияламайды, сондықтан цитокиннің синтезі жүрмейді.
Ил-5-тің стимульденген В- клеткалардың стимуляциясын күшейтеді және олардың бетінде ИЛ-2-ге арналған рецепторлардың экспрессиясын, IgA синтезін шақырады. Стимульденбеген В- клеткаларда ИЛ-5 IgM IgG секрециясын шақырады. Сонымен қатар ИЛ-5 эозинофилдерге хемоатрактант ретінде болады, паразитарлы инвазияларда олардың дегрануляциясын шақырады, атопиямен аллергиялық қабынудың патогенезіне қатысады. Ісікке қарсы активтілігі ИЛ-5-тің апаптозға қатысуымен байланысты. Жаңағы цитокиннің әсері Т-клеткаларға да тиеді. ИЛ-2 –нің қатысуымен ол цитотоксикалық Т- лимфоциттердің генерациясын шақырады.
Интерлейкин-6
Қарастырылып отырған цитокиннің көзі ретінде көптеген типіті клеткалар болады: Т-хлеперлер, моноцит- макрофагтар, фибробластар, эндотелиальді клеткалар, кератиноциттер. ИЛ-6-ның белсенді синтезі бактериялар, вирустар, митогендер, түрлі медиаторлар клеткаларға әсерінен кейін басталады. Көптеген эндогенді және экзогенді заттардың тобына жылдам және айқын реакциясы осы цитокиннің ерте медиаторлар тобына жататындығын көрсетеді. Мұндай қабілеттің бөтен патогеннің организмге түсуінде немесе тіндердің зақымдалу кезіндегі шұғыл реакцияның түзілуінде маңызы өте зор. ИЛ-6 ИЛ-1 мен TNF сияқты желел фазалар белоктар синтезін шақырады. Сондықтан қабынудың цитокиндер тобына жатады.
Ил-6-ның биологиялық әсерінің спектрі кең және нысана клеткалардың дифференцировкасын кеш кезеңдерінде қамтамасыз етеді. Негізінде В-клеткалардың дифференцировка факторы болып антидене түзуші клеткаларға дейін В-клеткалардың пісіп жетілуіне мүмкіндік жасайды. Костимулятор ретінде жаңағы цитокин митогенге немесе антигенге қарсы Т- клеткалардың пролиферативті жауабын дайындайды, антиген танушы Т- хелперлермен ИЛ-2-нің продукциясынның жоғарлауын қаматамасыз етеді.
ИЛ-6-ның жоғарлауы көптеген патологиялық жағдайларда анықталады: аутоиммунды ауруларда, псориазда, мезангиопролифративті гломерулонефритте, Капоши саркомасында, алкоголді циррозда, лимфомада, миеломада, бүйректің карциномасында. АИВ- инфицирленген адамдардың В-лимфоциттері TNF –а мен ИЛ-6 жоғары мөлшерде бөледі. Сонымен қатар атопиялық аллергиялық раекцияларда TNF-а мен ИЛ-6 жоғарлауы байқалған. ИЛ-6 деңгейнің жоғарлауы жаралы аурудың, панкреатитің, глютенді энреропатияның, Крон ауруының, арнайы емес калиттың, вирусты гепатитің, біріншілік билиарлы цирроздың асқынуында байқалады.
Интерлейкин-7
ИЛ-7 жетілмеген В- клеткалық ізашараларына пролиферативті әсер етеді: пре-В1 және пре-В2.
ИЛ-7-нің өсу реттеуші белсенділігі және сүйек кемігінің жетілмеген пре-Т- клеткаларымен тимусқа таралады.
ИЛ-7-нің басқа цитокиндерден ажырататын ерекшелігі- тіндік локализациясының шектелуі және бағыттау әсері болып табылады. Ил-7 ИЛ-2 мен бірге қатерлі ісіктердің иммунотерапиясында қолданылылуы мүмкін. ИЛ-7 ісікті клеткалардың апоптозын шақыруға қабілеті бар, жедел миелобластты лейкоз тобы клеткаларының дефференцировкасын шақырады.
Интерлейкин-8
Төмен малекулалы қабыну цитокині. Қабыну аймағына түрлі клеткалар типтерінің (нейтрофилдер, моноциттер, эозинофилдер, Т- клеткалар) хемотаксисын қамтамасыз етуші хемокиндер тобына жатады. Көбінесе цитокиндер (ИЛ-1 мен TNF) мен бактериалды эндотоксиндердің әсері арқылы бөлінеді.
ИЛ-8-дің продуцент клеткалары: макрофагтар, лимфоциттер, эпитемиальді клеткалар, фибробласттар, эпидермис клеткалары.
Басқа цитрокиндер сияқты, ИЛ-8- индуцибильді белок. Оның продукциясы митогендер немесе эндогенді регулятордың (ИЛ- 1,3, ФНО-, ГМ-КСФ және т.б.) клеткаларға әсерінен кейін басталады.
ИЛ-8-дің негізгі қызметі- нейтрофильдер, макрофагтар, лимфоциттер, эозинофилдер үшін хемоаттрактант ретінде қызмет ету. Осы биологиялық әсерінен басқа, ИЛ-8 нейтрофильдердің адгезивті қабілетін күшейтеді, интергриндер мен адгезивті қасиетті бар басқа қосылыстардың экспрессиясын өзгертеді. ИЛ-8-дің клеткалар миграциясын шақыру қасиеті патоген енген жерлерде жедел қабыну реакциясына белсенді түрде қатыстушы ретінде анықтайды.
Ил-8 жоғарлауы созылмалы немесе жедел қабыну жағдайлармен ассоциацияланады және жаралы колитпен қаттар ревматоидты артритте нейтрофилдердің тіндік инфильтрациясымен корреляциялайды. ИЛ-8 қабыну орнында ИЛ-1 мен TNF кейін пайда болады, псориазда маңызды орын алады.
Интерлейкин-10
Бұл лимфокин 17-21 kDa молекулярлы массамен, Т-клеткалармен (Тhy-2), бөлінетін бір қатарлы цитокиндердің антагонисті ретінде қаралуы мүмкін. ИЛ-10
Тhy-1 клеткаларымен бөлінетін INF-γ-ның продукциясын тәжейді. Сонымен қатар антигенмен митогендерге Т-клеткалардың пролифиративті жауабын және активтелген моноциттермен ИЛ-1 b, ТNF және ИЛ-6 секрециясын тежейді. Ил-10 IgE синтезін стимульдеуі мүмкін. Әртүрлі ісіктерде ИЛ-10-ның жоғарлауы анықталған. Сонымен қатар ИЛ-10 деңгейінің жоғарлауы нашар болжамдық көрсеткіші болып табылады.
Интерлейкин-11
ИЛ-11 сүйек кемігінің стромалды клеткаларымен синтезделеді. Нысана клекалары ретінде остеокласттардың гемопоэтикалық ізашарлары болады. Функциялық қасиеттері остеокласттердің түзілуі,қабынудың шақырушы цитокиндердің продукциясын тежеуі. ИЛ-11-ші ин виво және ин витрода Т-хелперлердің көмегімен антидене түзілуін күшейтеді. ИЛ-11 –ші мегакариоцитозды күшейтеді басқада қан клеткалардың дамуына әсер етеді, мысалы макрофагтардың. ИЛ-11-дің көзі ИЛ-1 мен стимульденген фибробластарда болады. ИЛ-1 және ИЛ-6 тәрізді жедел фазалы белоктардың синтезіне қатысады.
Интерлейкин-12
ИЛ-12 массасы 70 к Da гликопротеин болады, 2 гликолизирленген субъ бірліктен тұрады: р40 р35 дисульфидті байланыспен өзара байланысқан. Субъ бірліктер әр түрлі бір біріне тәуелсіз гендермен кодталады. ИЛ-12 бөлетін клеткалар көп мөлшерде р40-ты секреттейді. р40 рецептормен байланысуға қатысады, ал р35 сигналды өткізуге қажет. Ил-12-ші активтелген макрофагтармен бөлінеді және иммундық клеткалық реакцияларға әсер етеді. ИЛ-12-ші ЛАК жүйіенің клеткалардың литикалық активтілігін жоғарлатады. ИЛ-12-ші Т және NK клеткалардың активациясында өспелі фактор реттінде әсер етеді. Бұл жағдайда ИЛ-12 INF-γ секрециясының индукторы ретінде және ИЛ-4 пен индукцирленген IgE синтезінің ингибиторы ретінде әсер етеді.
ИЛ-12 мактофагтардың цитотоксикалық әсерін күшейтеді, ал оның макрофагтар арқылы продукциясының жетіспеушілігі ісікке қарсы активтілікті төмендетуге мүмкін. ИЛ-12 өкпенің қатерлі ісігінде ісікке қарсы әсерін көрсетеді. Тік ішек қатерлі ісігінің ұлғаюы ИЛ-12-нің продукциясының төмендеуімен және ИЛ-10-ның продукциясының күшеюімен ассоциациялайды.
Ісіктер некрозының факторы (ІНФ).
ІНФ-  және ІНФ-
және ІНФ- 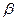
Тыныш күйдегі клеткалар- макрофагтар немесе лимфоциттер ІНФ-ға сәйкес продуцирленбейді. Олардың секрециясы тек индуктор әсерінен кейін басталады. Бұл қатынаста бактериялар мен олардың қабырғаларының компонеттері активті болып табылады.
Ісік клеткаларына ІНФ цитотоксикалық әсері ДНК деградациясы мен митохондри қызметінің бұзылуына байланысты. ІНФ литикалық әсері интерферон бар кезде күшейеді. Осы екі цитокиндердің сиергиялық әсер ету кезде күшейеді. Осы екі цитокиндердің синергиялық әсер ету мезханизмінің бірі- интерферон әсерімен ісік клеткаларына ІНФ рецепторларына экспессиясын күшейтуден тұрады.
Интерферондар тобы ИФН вирусқа қарсы белсенділік қабілетіне ғана емес (осы көрсеткіш бойынша олар ашылды), сонымен қатар иммунды жауапты реттеуде қатысады. Иммунды жүйе клеткаларының түзілуі мен миелопоэз үшін гранулоцитарлы- макрофагальді колониестимулирлеуші фактордың (ГМ-КСФ) маңызы зор.
В- клеткалармен және макрофагтармен өңделетін интерлейкин- 12 жуықта ашылғаны маңызды. Оның ТнО клеткаларын Т-қабыну клеткаларына (Тн1) индуцирлеу қабілеті иммунды жүйенің түзілуіне тікелей қатысы бар.
Қабыну реакцияларында, макрофагингиберлеуші фактордың (МИФ) белсенді ролі жатыр, ол қабыну аймағына тартып және сонда моноцит- макрофагтарды ұстап тұратын аттрактант ретінде қатысады.
Ең зерттелген кейбір цитокиндердің негізгі сипаттамасы
(қосымша мәліметтер)
Не нашли, что искали? Воспользуйтесь поиском: