ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Биологическая изменчивость. Исходный материал для селекции лесных древесных пород
1.4.1. Изменчивость живых организмов
Изменчивость — это процесс возникновения различий между особями по ряду признаков (размеры, форма, окраска, химический состав и пр.). Наряду с наследственностью и отбором она является основой органической эволюции. Более подробно явление изменчивости и методы ее изучения были уже рассмотрены при изучении курса генетики. Здесь необходимо вспомнить основные положения, которые характерны для изменчивости, поскольку этот параметр является одним из важнейших для эффективной селекции.
Выделяют изменчивость качественную (олигогенную, альтернативную), основанную на изменчивости по качественным признакам, и количественную (полигенную, непрерывную, флуктуирующую), основанную на изменчивости по количественным признакам.
Качественные признаки контролируются, как правило, одним или немногими генами, действие которых четко отделено от действия ненаследственных факторов. Различия по качественным признакам устанавливаются непосредственно путем наблюдения или сравнения без применения измерения или взвешивания. Качественные признаки обладают прерывистой изменчивостью. Описываются они по принципу “есть — нет”: опушенный — неопушенный, окрашенный — бесцветный, трещиноватый — гладкий, пирамидальный — раскидистый. Большая часть качественных признаков при гибридизации наследуется в соответствии с законами Г. Менделя.
Количественные признаки (мерные, полигенные) контролируются суммарным действием большого числа генов. Эти признаки могут быть определены путем измерения, взвешивания, подсчета. Вследствие полигенного контроля и большой модификационной изменчивости под влиянием условий внешней среды количественные признаки обладают непрерывной изменчивостью. Они могут быть описаны по принципу “больше — меньше”, т.е. один и тот же признак, присущий разным особям или формам, имеет различную степень выражения. К количественным признакам относятся: диаметр и высота ствола, прирост и запас, угол прикрепления сучьев, плотность древесины, урожайность семян и т.п. Проявление количественных признаков в значительной степени зависит от условий внешней среды.
Изменчивость органически присуща всем живым существам. Немецкие лесоводы еще в XIX веке констатировали, что у каждого дерева свое лицо. Ботаники попытались упорядочить результаты этой изменчивости. Так, Н.И. Вавилов, В.Н. Сукачев и др. вслед за исследованиями Ч. Дарвина, Э. Цедербауэра и голландского ботаника Я. П. Лотси показали, что виды слагаются из большого числа биотипов (форм), которые отличаются как морфологическими, так и физиологическими особенностями. Было установлено, что виды могут быть крупными или мелкими. Первые Лотси называл “ линнеонами ” в честь К. Линнея, а вторые — “ жорданонами ” в честь французского ботаника Жордана.
Среди систематиков до сих пор идут споры об объеме и отличительных признаках такой основной систематической категории, как вид. Однако постепенно на смену представлениям о неделимом монотипном виде приходит понятие о политипической структуре вида, сторонниками которой были и наши ученые В.Н. Сукачев и Н.И. Вавилов.
В.Н. Сукачев отмечал среди внутривидовых следующие категории:
• экотипы — наборы биотипов, отвечающие определенным условиям существования, включающие климатипы, эдафотипы, ценотипы — как результат групповой изменчивости;
• лузусы — формы, отличающиеся какими-либо признаками, но не обладающие определенным ареалом или не относящиеся к определенным экотипам, например красно- и зелено-шишечные формы ели;
• аберрации — редкие резко уклоняющиеся формы: змеевидные, плакучие и т. п.;
• экады — ненаследственные формы, или модификации.
Для практической селекции очень большое значение имеет закон гомологических рядов в наследственной изменчивости, сформулированный Н.И. Вавиловым в 20-е годы и доложенный в Саратове на III Всероссийском съезде селекционеров в июне 1920 г. Сущность закона заключается в том, что признаки близкородственных видов и даже родов изменяются параллельно. Например, плакучие и пирамидальные формы есть и у тополя черного, и у тополя белого. Более того, и у рода тополя в целом, и у рода ивы, и у других родов древесных растений. Таким образом, зная формовое разнообразие одного вида или рода, можно прогнозировать его у родственных видов.
В последние десятилетия изменчивость лесных пород стали изучать на биохимическом уровне. При этом исследуются изозимы (аллозимы), антигены, терпены и другие вещества.
Изозимы — это множественные формы энзимов (специализированных белков, которые ускоряют биохимические реакции), синтез которых контролируется более чем одним геном, в то время как аллозимы — альтернативные формы фермента, кодируемые различными аллелями одного и того же гена. К настоящему времени для самой изученной в этом плане сосны обыкновенной исследовано 26 систем энзимов, для дуба — немногим более 10. Определение энзимов производят в крахмальном геле с помощью электрофореза. Аллели обозначаются номерами от катода к аноду.
Как и все биохимические исследования, эти методы требуют узкой специализации персонала, недешевых лабораторного оснащения и расходных материалов. Поэтому они могут быть использованы только в немногих научных организациях.
Однако эти методы не всегда отражают реально существующее разнообразие и изменчивость признаков. Так, А. И. Ирошников (1996) отмечает контрастность характеристик межпопуляционной изменчивости видов с обширными ареалами (сосен обыкновенной и сибирской, лиственницы сибирской), полученных по материалам изоферментного анализа, с одной стороны, и результатами испытания потомств популяций в географических культурах, созданных в разных регионах, с другой стороны. Если в первом случае фиксируется как правило 3-5%-ный (0,5-12%-ный) уровень межпопуляционной генетической изменчивости, то во втором — он достигает 20-30% и более.
В общем виде изменчивость можно выразить следующей формулой:

где 
 — наблюдаемая фенотипическая изменчивость;
— наблюдаемая фенотипическая изменчивость; 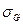 — генотипическая изменчивость;
— генотипическая изменчивость;  — изменчивость, обусловленная средой;
— изменчивость, обусловленная средой; 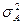 — аддитивная доля изменчивости в общей фенотипической изменчивости;
— аддитивная доля изменчивости в общей фенотипической изменчивости;
 — неаддитивная доля изменчивости;
— неаддитивная доля изменчивости;  — доля изменчивости, обусловленная доминированием;
— доля изменчивости, обусловленная доминированием;  —доля изменчивости, обусловленнаяэпистазом.
—доля изменчивости, обусловленнаяэпистазом.
Кроме того, фенотипическая изменчивость некоторыми исследователями (С. А. Мамаев) делится на: внутривидовую и внутриорганизменную, или эндогенную.
Внутривидовая изменчивость подразделяется на:
• индивидуальную — проявление дифференциации особей в пределах вида;
• половую — зависящую от пола;
• хронографическую (временную), включающую возрастную (онтогенетическую) и сезонную изменчивости;
• экологическую, обусловленную воздействием нарастениеопределенных факторов окружающей среды;
• географическую — результат дифференциации вида в пределах ареала в широтном и меридиональном направлениях;
• гибридогенную — на границах ареалов в местах спонтанной гибридизации.
Эндогенная (или метамерная) изменчивость — это изменчивость органов (листьев, цветков, плодов, семян и т. п.) в пределах особи. В зависимости от уровня изменчивости тех или иных видов оценивается их исходный материал. Изучение изменчивости исходных популяций позволяет выбрать наиболее эффективный метод селекции.
1.4.2. Исходный материал для селекции лесных древесных пород
Под исходным материалом подразумевают культурные и дикие формы растений, используемые для выведения новых сортов. Основными источниками исходного материала являются естественные (местные и интродуцированные) популяции, гибридные популяции, полученные на основе внутривидовых и отдаленных скрещиваний, инцухт-линии, индуцированные мутанты и полиплоиды (Г.В. Гуляев, В.В. Мальченко, 1983).
Успех селекции обусловлен формовым разнообразием древесных растений. Лесные деревья настолько изменчивы, что почти невозможно найти два экземпляра, которые были бы полностью сходны по строению, внешнему виду и жизненным проявлениям (Э. Ромедер, Г. Шенбах, 1962). Чтобы ориентироваться в этом многообразии, ботаники делят растительный мир на классы, семейства, роды, виды, подвиды, разновидности и другие надвидовые и внутривидовые таксоны.
Причиной фенотипического разнообразия являются внешние (среда) и внутренние (наследственность) факторы. Разграничить влияние среды и наследственности на формирование конкретного фенотипа возможно только при постановке специальных опытов, в которых нормируется влияние среды или генотипа. Сила этих влияний могут быть самыми разными в зависимости от условий испытания, характера исследуемого материала и изменчивости изучаемых признаков. При этом различают изменчивость морфологических (форма ствола, кроны, корки; форма, длина, ширина, толщина и окраска хвои и листьев; цвет и форма цветков, шишек, семян, и т.п.), физиологических (фенологические признаки, рост и др.), качественных признаков, а также признаков устойчивости.
Многообразие форм является результатом непрерывного мутационного процесса, отдаленного внутри- и межвидового скрещивания и полиплоидизации. Степень полиморфизма зависит от возраста видов, а также условий их произрастания. Филогенетически молодые и эволюционно старые виды характеризуются, как правило, меньшей внутрипопуляционной изменчивостью по сравнению с видами, находящимися в расцвете своего развития. Гетерогенность популяций обычно значительна в области оптимума вида. В экстремальных условиях изменчивость деревьев по ряду признаков выражена слабее, наряду с этим частота отдельных форм здесь бывает повышенной.
За исключением случаев модификаций, связанных с влиянием биотических и абиотических факторов, полиморфизм популяций генетически обусловлен. Однако имеются значительные различия в степени генетического контроля тех или иных признаков. При этом выделяется группа константных, строго детерминированных признаков, не зависящих от внешних факторов, и группа лабильных признаков, изменяющихся в той или иной мере под влиянием среды. Характер изменчивости некоторых признаков у древесных растений зависит также от типа их онтогенеза.
Фенотипическое проявление нормы реакции особей в каждом случае находится в определенных пределах, что связано с функциями соответствующих органов, обеспечивающих индивидууму жизне- и конкурентоспособность в довольно широких диапазонах среды произрастания. В изменении отдельных групп признаков наблюдаются корреляции. Последние отмечаются, в частности, между некоторыми морфологическими признаками древесных растений и физико-техническими свойствами древесины, интенсивностью их роста, смолопродуктивностью, устойчивостью к биотическим и абиотическим факторам и другим свойствам, имеющим важное хозяйственное значение.
Многообразие форм древесных пород давно было подмечено ботаниками, садоводами и лесозаготовителями. На первом этапе дендрологов прежде всего интересовали резко уклоняющиеся формы с декоративными свойствами, представляющие особую ценность для зеленого строительства. В течение XVII-XIX веков в парках и ботанических садах многих стран мира коллекционировали деревья с пирамидальной, плакучей, колонновидной, зонтикообразной, шаровидной, веерообразной, шатровидной как плотной, так и ажурной формами кроны;
с причудливыми ветвлениями и изгибами стволов; разнообразными наростами и наплывами на ветвях и стволе;
гладкой, отслаивающейся или мощной коркой;
с признаками гигантизма или, наоборот, карликовости;
с кустовидной или стелющейся формой;
с различными окраской, деформацией (рассеченность, скрученность) и опущением листьев;
с яркой окраской или аномалиями в развитии, размере и форме цветков, микро- и мегастробилов, плодов, шишек и др. Изучение формового разнообразия стимулировали и работы Н.И. Вавилова по параллельной изменчивости у древесных растений.
Из всего многообразия селекционеры отбирают ценные в каком-либо отношении фенотипы и разрабатывают меры по их сохранению. При этом предлагается:
а) выделение лесных генетических резерватов;
б) сохранение отдельных площадей (в особенности изолятов) насаждений и деревьев (эталонных, плюсовых);
в) создание испытательных (коллекционных) культур и кленовых архивов;
г) создание банков семян, пыльцы и меристем (С.А. Мамаев и др., 1988). Более подробно проблема сохранения генетического фонда лесных древесных пород в местах естественного (in situ) и вне мест естественного (ex situ) произрастания (плантации, коллекции, архивы, банки семян, меристем, пыльцы и т.п.) рассмотрена ранее.
К сожалению, в силу ряда причин не все из полученных и отмеченных в настоящей разделе достижений нашли использование в производстве. Кроме того, в последние годы многие из направлений исследований пришлось свернуть из-за недостатка средств. Наблюдаются неблагоприятные тенденции и в других странах мира. Они заключаются в истощении невозобновляемых ресурсов, уменьшении биологического разнообразия, снижении доли лесопокрытой площади, вырубке наиболее производительных насаждений, развитии других неблагоприятных факторов, с одной стороны, и увеличении потребности в различных видах полезностей леса, с другой. Очевидно, что требуются какие-то меры по преодолению этих неблагоприятных тенденций. Необходимо наметить задачи ближайшего будущего (программа-минимум) и более отдаленного (программа-максимум).
Программа-максимум должна базироваться на главной идее отечественного лесоводства — неистощительности пользования, а также на идее западного мира об устойчивом лесоуправлении (sustainable forestry}. Она должна включать не только оптимальные технологии рубок и лесовосстановления, но и защиту ценного генофонда лесных древесных пород, а также выведение новых генотипов и сортов для его обогащения.
Программа-минимум должна содержать практические рекомендации по ведению лесного хозяйства на ближайшие 10-20 лет, то есть показать, что можно сделать при объективном наличии вышеуказанных тенденций и сегодняшних реальных возможностей. Среди других приоритетов она должна включать углубление образования в области лесной генетики и селекции, сохранение ценного природного генофонда лесных древесных пород и долговременных производственных и опытных объектов, развитие теоретических и малотрудоемких прикладных исследований, обобщение имеющегося опыта и научных результатов в области лесной генетики и селекции.
Реализация таких подходов позволит повысить продуктивность создаваемых насаждений и сохранить генетический потенциал лесных древесных пород для наших потомков.
Вопросы для самопроверки
1. Дайте определение селекции как науки. Назовите основные разделы селекции, которые сформулировал Н.И. Вавилов.
2. Приведите задачи селекции лесных древесных пород в трудах русских ученых С.З. Курдиани, Н.С. Нестерова,Г.Ф. Морозова и практические результаты в этой области, полученные в канун первой мировой войны.
3. Назовите основные периоды развития селекции лесных древесных пород у нас в стране после первой мировой войны.
4. Осветите основные направления развития отечественной селекции лесных древесных пород в 1921-1932 гг.
5. Покажите направления исследований и результаты по селекции лесных древесных пород в 1933-1953 гг.
6. Раскройте содержание основных направлений исследований и достигнутых результатов в 1953-1967 гг. и в последующие годы.
7. В чем заключаются программные подходы в селекции лесных древесных пород?
8. Охарактеризуйте проблему изменчивости живых организмов. Покажите вклад в исследование проблемы Н. И. Вавилова, В. Н. Сукачева, С. А. Мамаева.
9. Приведите краткую характеристику биохимических методов исследования разнообразия лесных древесных пород.
10. Приведите формализованное представление об изменчивости в математическом выражении.
11. Что такое исходный материал для селекции? Приведите примеры многообразия форм у различных лесных древесных пород.
Глава 2.
Не нашли, что искали? Воспользуйтесь поиском: