ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
Джгутування у бактерій
 |  |  |  | ||||
Монотрих (а) Лофотрих (б) Амфітрих Перитрих (в)
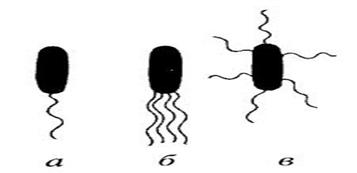
Рис. 3.29 Види розміщення джгутиків у бактерій
За допомогою джгутиків бактерії здійснюють спрямований рух (таксис), який визначається зовнішніми стимуляторами. Таксис може бути як позитивним, так і негативним. Причинами такої реакції можуть бути:
· різниця в концентрації хімічних речовин в середовищі (хемотаксис)4
· різниця в концентрації кисню (аеротаксис);
· різниця в інтенсивності освітлення (фототаксис);
· оріентація в магнітному полі (магнітотаксис).
Побачити джгутики можна в електронному мікроскопі після негативного контрастування або обробки препарату солями важких металів, в світловому – тільки після використання спеціальних методів фарбування.
Звивисті бактерії (спірохети) рухаються не за допомогою джгутиків, а інших органоїдів – аксіальних фібріл (від 2 до 100 штук на клітину). Ці нитковидні структури закріплені на полюсах спірохети, і 2 пучка перекриваються в центральній частині клітини (рис.3.30).

Рис.3.30 Аксиальні фібріли спірохет
Обертання фібрил викликає еластичну хвилю на поверхні спірохети, і остання також обертається, вигинається, рухається хвильо- або гвинтоподібно.
Фімбрії та пілі – поверхневі структури багатьох бактеріальних клітин, які мають вигляд білкових циліндрів довжиною 1-1,5 мкм та діаметром 7-10 нм. Розрізняються за будовою та призначенням, при цьому у одної клітини можуть бути присутні вирости 2-х типів (табл. 6, рис.3.31).
Табл.6
Характеристика білкових виростів
| Назва органоїду | Хімічний склад, кількість | Розміри та розміщення | Функція |
| Фімбрії | Білок пілін. (від 1 до декількох тисяч) | Довжина 0,3-0,5 мкм,діаметр 5-10 нм По всій поверхні клітини. | З’єднання клітин при утворенні плівок, прикріплення до субстрату. Такі структури мають бактерії будь-якої форми. |
| Пілі | Білок пілін. (від 1 до 5) | Довжина 1-2 мкм, діаметр 8-10 нм. В будь-якому місці на поверхні клітини | Перенесення генетичного матеріалу прокаріотів (маленькі, додаткові кільцеві молекули ДНК – плазміди) в сусідні клітини. |

Рис. 3.31 Фімбрії та пілі у бактерій
Фімбріїї – коротенькі багато чисельні вирости, наявні у всіх мікробних клітин. Стійкі до дії певних хімічних речовин (6 М сечовина, 1М NаОН, додецилсульфат натрію, трипсин), руйнуються при кип᾽ятінні в розчинах з кислим рН. Розміщені по всій поверхні клітини (50-400 шт. діаметром 7 нм).
Фімбріїї приймають участь в прикріпленні клітини до різноманітних субстратів (висока адгезивність кінцевих ділянок). Забезпечують приєднання до органічних та неорганічних частинок, еритроцитів, інших тваринних клітин та мікроорганізмів. Клітини з такими пілями отримують переваги при вирощуванні і на твердих середовищах, і в рідкому середовищі (утворення плівки).
Пілі – це більш довгі і нечисленні вирости (1-5 шт.), які приймають участь у так званих парасексуальних процесах. Такі вирості представляють собою білкові циліндри, перпендикулярні поверхні клітини діаметром 9-11 нм. Для їх утворення необхідна активність 13 генів.
Статеві пілі виникають у тих клітин, які мають в цитоплазмі коротенькі кільцеві молекули ДНК (плазміди). Їх утворюють клітини в активній фазі росту (ентеробактерії, псевдомонади, вібріони, пастерели тощо). Генетичний матеріал плазмід різних типів визначає форму та довжину піля, його призначення та місце прикріплення до оболонки сусідньої клітини.
Пілі легко від’єднуються від клітин під дією органічних розчинників. У клітін, які примусово втратили пілі, швидко утворюються нові. За 30 секунд піль досягає 1/2 нормальної довжини, а повністю формується через за 4-5 хвилин. Сформовані піли існують на поверхні клітин зберігаються на поверхні мікробів впродовж 4-5 хвилин, а потім - відкидаються.
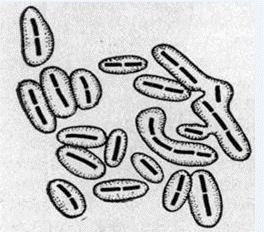
Спеціальні зовнішні і внутрішні структури бактерій (КАПСУЛИ, ЕНДОСПОРИ)
Капсула -це слизова оболонка на поверхні бактеріальної клітини (рис.3.32). Основна функція даного органоїду – захисна. Капсула захищає бактеріальну клітину від дії несприятливих факторів оточуючого середовища (висихання, дія фагів, наявність запасних поживних речовин та ін.). Але при вилученні слизового шару клітина продовжує рости та розмножуватись

а б
Рис.3.32 Капсули бактерій: а – розміщення;
б – види капсул (№3 - мікрокапсула, №4 - макрокапсула, №5 - слизовий шар).
Капсула складається з шарів фібрил, які розміщені паралельно або перпендикулярно клітинній стінці. Основна речовина – поліцукри (D-галактоза, D-маноза, D-глюкуронова кислота, L-рамноза). Капсули стрептококів та бацил мають ще поліпептиди, побудовані із залишків D- і L-глутамінової кислоти). Наявність капсули залежить від штаму мікроорганізму (у одного виду існують капсульні та безкапсульні клітини), віку культури та умов культивування.
Відповідно до розмірів слизового шару виділяють:
· мікрокапсулу (товщина 0,2 мкм, реєструється за допомогою електронного мікроскопу);
· макрокапсулу (товщина – більша, ніж 0,2 мкм);
· слиз (товщина шару – більша за діаметр клітини).
Ендоспори бактерій. У бактерій певних родів в несприятливих для росту і розмноження умовах всередині клітини утворюється спеціальна структура (ендоспора), а в життєвому циклі бактеріальної клітини настаєстадія спокою.
Такими умовами є:
- нестача карбону, нітрогену, фосфору та ін. елементів в живильному середовищі;
- накопичення в середовищі катіонів калію та мангану;
- зміни рН та кількості кисню в середовищі;
- високий рівень опромінення середовища існування мікробів.
Від вегетативної клітини ендоспора відрізняється: мінімальною кількістю води; наявністю сполук, які не входили до складу вегетативної клітини; припиненням обміну речовин та процесу подвоєння генетичного матеріалу клітини. Основна функція спори – збереження виду в несприятливих умовах. Зрілі спори зберігаються в навколишньому середовищі впродовж сотен і навіть тисяч років, здатні витримувати високі (‹ 120 оС) та низьки температури, опромінення, висушування, дію токсичних речовин та ін.
Спороутворення характерно для 15 родів бактерій (Вacillus, Clostridium, Sporosarcina, Spirillum, Desulfotomaculum). Найкраще цей процес вивчено у паличковидних бактерій, які належать до родів Вacillus та Сlostridium.
Розміри спор 0,8-1,0 на 1,2-1,5 мкм, форма – сферична або овальна. Будова зрілої спори у різних видів бактерій практично однакова (рис.3.33).
Процес спороутворення відбувається в декілька етапів, триває 6-7 годин від початку дії несприятливого фактору, контролюється багатьма генами (›100 шт.). Комплекс генів має назву спорулон. У багатьох бацил під час утворення спор синтезуються антибіотики білкової природи, які припиняють ріст власних вегетативних клітин.
1 етап. Припиняється процес подвоєння ДНК та ріст клітини В клітинній стінці починається утворення сполуки, характерної тільки для спори (діпіколінова кислота). Ця речовина, у вигляді її кальцієвої солі, складає 10-15% від сухої маси зрілих спор.
2 етап. Цитоплазматична мембрана вегетативної клітини впячується від періферії до центру і відділяє нуклкеїд з ділянкою ущільненої цитоплазми від частини клітини. Через деякий час мембрана обростає цю зону і всередині материнської клітини утворюється проспора. Навкруги останьої починають формуватись шари муреїну, зовнішня частина яких перетворюється на кортекс (муреїн кортексу більш кислий у порівнянні з муреєном клітинної стінки). Надалі відбувається формування багатошарових покривів.
3 етап. Спора набуває характерної форми і займає в клітині певне положення. С початку дії несприятливого фактору до утворення зрілої спори проходить приблизно 6 годин.

Рис.3.33 Будова зрілої спори.
Зріла ендоспора являє собою зневоднене багатошарове утворення:
1. серцевина спори, спороплазма або кор містить рибосоми, ферменти, ліпіди та ін. речовини. В центральній частині знаходяться білки та нуклеїнові кислоти (50-60% від сухої маси спори);
2. цитоплазматична мембрана та залишки клітинної стінки;
3. кортекс (13-60% від об’єму спори);
5. зовнішня мембрана;
6. трьохшаровий покрив, складений з білків з незначною кількістю вуглеводів та ліпідів;
7. екзоспориум (білки, вуглеводи, ліпіди). Максимальна кількість шарів – до 15, товщина більше 100 нм.
Спори в клітинах бацил розміщуються за такими варіантами:
· у мікробів роду Вacillus ендоспора знаходиться в центрі (бацилярне або центральне положення);
· у мікробів роду Сlostridium вона зсунута від центру до полюсу (клострідіальне або субтермінальне положення) або знаходиться на одному з полюсів (плектридіальне або термінальне положення).

Рис. 3.34 Розміщення ендоспор у паличковидних бактерій певнх родів: бацилярне (1,4), клостридіальне (2,3,5).
Мікроорганізми роду Вacillus (В. anthracis, В. cereus та ін.) одночасно із спорами утворюють параспорові тільця. Це можуть бути як білкові кристали (В. thuringiensis), так і сферичні утворення діаметром 120-200 нм (В. anthracis), розміщені ізольовано або на поверхні спор.
Процес проростання спори починається із всмоктування води в присутності глюкози та деяких амінокислот, які прискорюють повернення спори до стану вегетативної клітини. Відбувається оводнення всіх структур спори та активація ферментів (літичні – руйнують багатошарові покриви). В середовище викидається діпиколінова кислота та іони кальцію. Оболонки розриваються, утворюється ростова трубка нової клітини. Час, за який спора перетворюється на вегетативну клітину займає 3-5 годин.
ЛІТЕРАТУРА
1. Ветеринарная микробиология и иммунология / Н.А.Радчук, Г.В. Дунаев, Н.М. Колычев и др.: Под ред. Н.А.Радчука. – М.: Агропромиздат, 1991. – 383 с.
2. Лукомская К.А. Микробиология с основами вирусологии – М.: Просвещение, 1987. – 192 с.
3. Теппер Е.З., Шильникова В.К., Переверзева Г.И. Практикум по микробиологии– М.: Колос, 1979. – 216 с.
4. Кузьмин В.В. Ветеринарная микробиология– М.: Сельхозгиз, 1958. – 231 с.
5. Сєги Й. Методы почвенной микробиологии / Пер. с венг.: Под ред. Г.С.Муромцева– М.: Колос, 1983. – 296 с.
Не нашли, что искали? Воспользуйтесь поиском: