ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
B- Окиснення вищих жирних кислот
Теорія окислення вищих жирних кислот була висунута А. Кноопом у 1904 р. Він висловив припущення, що окиснювання молекул жирної кислоти в тканинах організму відбувається в b-положенні шляхом послідовного відщіплення від молекули жирної кислоти з боку карбоксильної групи двох карбогенових фрагментів. У зв'язку з цим процес окислення жирних кислот називають b-окисленням.
Оскільки будь-яка жирна кислота, що входить до складу природних жирів, має парне число вуглецевих атомів, то відщіплення в кожній реакції по парі карбогенів обов'язково приводить процес до утворення масляної кислоти. Після чергового циклу b-окислення вона перетворюється в ацетооцтову кислоту, що гідролізується до двох молекул оцтової кислоти. Ця теорія не втратила свого значення дотепер і є основою сучасних уявлень про механізм окиснювання вищих жирних кислот.
b-Окислення вищих жирних кислот відбувається в мітохондріях клітин при участі мультиферментного комплексу.
Початковим етапом b-окислення є активація молекул вищих жирних кислот при участі АТФ і HSKoА. При цьому утворюються активовані форми: КоА-ефіри цих кислот.
Утворюється активна форма (ацетил-КоА) з жирної кислоти і коензиму А за рахунок енергії АТФ за участю ферменту ацетил-КоА-синтетази. Перенос ацильних груп з цитоплазми на матрикс мітохондрії здійснює карнітин. При участі ферменту карнітин-ацил-трансферази синтезується ацилкарнітин, здатний проникати усередину мітохондрій. Після чого карнітин повертається в цитоплазму клітки, а ацетил-КоА піддається окисленню.

Наступний етап - дегідрування жирних кислот за участю ФАД, при цьому відщепляються два атоми гідрогену в a- і b-положеннях і ацил-КоА перетворюється в КоА-ефир ненасиченої кислоти (єноїл-КоА).
На стадії гідратації єноїл-КоА, взаємодіючи з молекулою води, перетворюється в b-оксикислоту (b-гідрооксиацил).
Друга стадія дегідрування протікає при участі НАД - залежної дегідрогенази з утворенням b-кетокислоти (b-кетоацил-КоА).

Потім відбувається тіолазна реакція: взаємодія кетоацил-КоА з ацетил-КоА.

У результаті цієї реакції внаслідок розщеплення b-кетоацилу виділяється укорочена на два карбогенових атоми активована вища жирна кислота і двовуглецевий фрагмент у виді ацетил-КоА. Ацетил-КоА, що утворився, окислюється в циклі трикарбонових кислот, а активована вища жирна кислота багаторазово проходить весь шлях b-окиснення аж до утворення чотирьохвуглецевої сполуки - бутирил-КоА, що у свою чергу окисляється до двох молекул ацетил-КоА.
Енергетичний ефект b-окиснення. Число циклів окислення, яким піддається вища жирна кислота, залежить від кількості карбогенових атомів у її молекулі. При окисленні однієї молекули жирної кислоти утвориться n/2 молекул ацетил-КоА, де n - кількість атомів карбогену, а цикл повториться (n/2 - 1) раз, тому що молекула бутирил-КоА відразу розщеплюється на дві молекули ацетил-КоА. У кожнім циклі з'являються молекула ФАДН2 і молекула НАДН2. Молекула ФАДН2 при окиснюванні в дихальному ланцюзі і сполученого з ним фосфорилювання дає дві молекули АТФ, а НАДН2 - три молекули АТФ, тобто за один цикл b-окиснювання утворюється п'ять молекул АТФ.
Кожна молекула ацетил-КоА включається в цикл трикарбонових кислот, поступово розщеплюється до СО2 і Н2О з виділенням 12 молекул АТФ. Як приклад розглянемо b-окиснення пальмітинової кислоти.
При окисненні пальмітинової кислоти відбувається сім циклів b-окиснювання - (16/2 - 1), що веде до утворення 35 молекул АТФ. У результаті b-окиснення цієї кислоти утворюється вісім молекул ацетил-КоА (16/2), кожна з яких, окисляючись в циклі трикарбонових кислот, дає 12 молекул АТФ, тобто утворює 96 молекул АТФ. Таким чином, сумарний вихід енергії при окисленні однієї молекули пальмітинової кислоти складе: 35 + 96 = 131 молекула АТФ. Оскільки одна молекула АТФ була витрачена на активізацію вищої жирної кислоти на початку процесу вихід енергії складе 130 молекул АТФ. Близько 45 % усієї потенційної енергії пальмітинової кислоти при її окисленні в організмі може бути використана для ресинтезу АТФ, інша утилізується у виді теплоти.
Окиснення ненасичених жирних кислот відбувається так само, як і насичених, але має свої особливості, обумовлені положенням подвійних зв'язків. До початку b-окиснення в молекулі жирної кислоти відбувається переміщення подвійного зв'язку з положення 3-4 у 2-3 і зміна конфігурації подвійного зв'язку з цис- у транс-положення.
Більшість природних ліпідів містить жирні кислоти з парним числом вуглецевих атомів. Однак у ліпідах рослин і деяких морських організмів виявляються жирні кислоти з непарним числом вуглецевих атомів. Вони також піддаються b-окисненню, у результаті якого з'являються ацетил-КоА і пропіоніл-КоА. Останній перетворюється в сукциніл-КоА - метаболіт циклу Кребса.
Процес b-окиснення вищих жирних кислот за участю НS коензиму А активніше протікає в печінці, жировій тканині, серцевому і кістяковому м'язах, слабкіше - у нирках, підшлунковій залозі та інших органах.
Метаболізм кетонових (ацетонових) тіл
Кетоновими (ацетоновими) тілами називають ацетооцтову кислоту (ацетоацетат) СН3-СО-СН2-СООН, b-гідрооксимасляну кислоту (b-оксибутират) СН3-СНОН-СН2-СООН і ацетон СН3-СО-СН3. Вони утворюються в печінці з ацетил-КоА. На першій стадії цього процесу з двох молекул ацетил-КоА утворюється ацетоацетил-КоА.

~
Потім ацетоацетил-КоА взаємодіє ще з одною молекулою ацетил-КоА.

b-гідрокси-b-метилглутарил-КоА, що утворився, розщеплюється на ацетоацетат і ацетил-КоА.
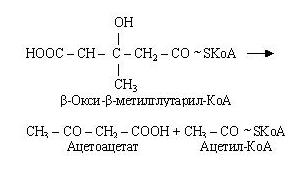
Ацетоацетат (ацетооцтова кислота) при участі НАДН2 відновлюється до b-гідромасляної кислоти (b-гідрооксибутират).

Крім того, ацетоацетат при декарбоксилюванні перетворюється в ацетон. Ацетонові тіла розглядаються як специфічні, а не проміжні продукти обміну біосинтезу жирних кислот.
В крові здорової людини кетонові тіла містяться лише в невеликих кількостях.
При важких формах діабету, повному голодуванні, недостатньому змісті вуглеводів у їжі концентрація кетонових тіл у крові збільшується і може досягати 20 ммоль/л, розвивається ацидоз (кетоз), при цьому швидкість утворення кетонових тіл перевищує здатність периферичних тканин їх утилізувати.
В даний час доведена важлива роль кетонових тіл у підтримці енергетичного гомеостазу. Вони є джерелами енергії для роботи м'язів, нирок і діють як частина регуляторного механізму зі зворотним зв'язком, запобігаючи надзвичайній мобілізації жирних кислот з "жирових депо". Отже, розвиток гіперліпемії сприяє заощадженню вуглеводів при їхньому недоліку. Виключенням є печінка і головний мозок, що не використовують кетонові тіла як джерело енергетичного матеріалу. У м'язах і інших тканинах ацетонові тіла окисляються з утворенням АТФ.
12.3. БІОСИНТЕЗ ЛІПІДІВ
Здатність людини і тварин запасати глікоген обмежена, тому кількість глюкози, що надходить в організм, може перевищувати його енергетичні потреби. При цьому глюкоза використовується в якості "будівельного матеріалу" для синтезу жирних кислот і гліцеролу, що перетворюються в триацилгліцерол, що відкладається в "жирових депо".
У синтезі триацилгліцеролів умовно виділяють три етапи: утворення гліцеролу, синтез жирних кислот, взаємодія гліцеролу і вищих жирних кислот з одержанням складних ефірів.
Гліцерол у виді гліцерофосфату утвориться або з продукту розпаду вуглеводів 3-фосфо-гліцеролового альдегіду, або з гліцерофосфату, що виділяється при розщепленні триацилгліцеролів у клітках тканин.
Біосинтез жирних кислот полягає в поступовому подовженні ланцюга жирної кислоти на два вуглецевих атоми до виникнення визначеної жирної кислоти. У цьому процесі беруть участь ацетил-КоА, малоніл-КоА, АТФ, НАДФН2, ацилпереносячий білок і система ферментів, з’єднаних у мультиферментний комплекс - синтетаза жирних кислот.
"Стартовою" речовиною біосинтезу жирних кислот є малоніл-КоА. Він утворюється з ацетил-КоА шляхом приєднання активованого оксиду вуглецю.

Синтетаза складається із семи ферментів, що утворюють сферичну частку, у центрі якої розташований ацетилпереносячий білок (АПБ). До нього при синтезі приєднуються вихідні субстрати - ацетил-КоА, малоніл-КоА, і він переміщає їх від одного ферменту до іншого.
На початку синтезу відбувається приєднання ацетил-КоА і малоніл-КоА до синтетази при участі АПБ і утворюється ацетоацетил-АПБ. Потім він переміщається до наступного ферменту (редуктази), що передає йому два атоми гідрогену від НАДФ·Н2. Речовина, що утворюється, взаємодіє з ферментами, що входять до складу синтетази. Цим закінчується один цикл синтезу, і карбогеновий ланцюг кислоти подовжується на два карбогенових атоми.
Не відщеплюючись від АПБ, жирна кислота включається в наступний цикл синтезу, що починається з приєднання до неї малоніл-КоА. Цей процес повторюється доти, поки не синтезується потрібна для клітини жирна кислота з визначеною кількістю карбогенових атомів. Кислота, що утворилася, відщеплюється від синтетази, вступає в інші реакції, а до синтетази приєднуються вихідні субстрати, і починається синтез наступної молекули насиченої жирної кислоти.
Ненасичені жирні кислоти в організмі людини не синтезуються, тому вони повинні надходити з їжею.
На останньому етапі синтезу триацилгліцеролів відбувається взаємодія гліцерофосфату з двома молекулами активованих жирних кислот. Спочатку утворюється фосфатидна кислота, що є проміжним продуктом при утворенні триацилгліцеролів і фосфоліпідів.
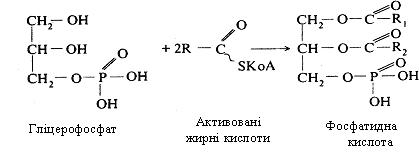
Потім від цієї кислоти відщеплюється фосфорна кислота, до диацилгліцеролу, приєднується ще одна молекула активованої жирної кислоти і утворюється триацилгліцерол (рис. 12.2.).

Рис 12.2. Синтез тригліцеролів
У процесі синтезу фосфоліпідів з'являються диацилгліцерол і активна форма холіну - цитідиндифосфохолін (ЦДФ-холін), що взаємодіють між собою з утворенням лецитину. Найбільше активно цей процес відбувається в печінці, з якої вони розносяться до органів і тканин (рис. 12.3.).

Рис. 12.3. Синтез фосфатидилхолінів.
12.4. РЕГУЛЯЦІЯ ОБМІНУ ЛІПІДІВ І ЙОГО ПОРУШЕННЯ
Обмін ліпідів в організмі залежить від впливу ряду факторів внутрішнього і зовнішнього середовища. Істотну роль грають також вік, стать, характер харчування, вид трудової діяльності, режим дня, форми відпочинку, кліматогеографічні умови проживання і т.д. Активує процеси синтезу ліпідів і призводить до надлишкового їх накопичення в "жирових депо", а надалі до порушення обміну і виникненню захворювань, такий фактор, як нерегулярне харчування, особливо якщо велика калорійність раціону приходиться на другу половину дня і вечірні години. Несприятливо впливає на обмін жирів різка перевага в раціоні засвоюваних вуглеводів, моносахаридів, що утворюються з них, при малорухомому способі життя повною мірою не використовуються, а перетворюються в жири.
Ліпідний обмін в організмі регулюється центральною нервовою системою. Кора головного мозку впливає на жирову тканину через симпатичну і парасимпатичну нервову систему і ендокринні залози. Кількість жиру в "жирових депо" зменшується при тривалому негативному емоційному стресі, що супроводжується збільшенням викиду гормону надниркових залоз адреналіну в кровоносне русло, що призводить до зменшення маси тіла. Цей ефект пояснюється тим, що жирова тканина рясно інервована волокнами симпатичної нервової системи, а норадреналін, що виділяється, як і адреналін, збільшує швидкість ліполізу в жировій тканині. Крім того, адреналін через систему відповідних ферментів сприяє утворенню активної форми ліпази. Дія глюкагону і тироксину подібно впливу адреналіну і норадреналіну (катехоламінів): вони стимулюють ліполіз.
На ліпідний обмін впливає гормон росту (СТГ), що утворюється в передній частці гіпофіза. При недоліку цього гормону збільшується відкладення жиру в організмі, розвивається гіпофізарне ожиріння. При підвищеній продукції його збільшується ліполіз і вміст жирних кислот у плазмі крові.
Інсулін має дію, протилежну адреналінові і глюкагону: він зменшує утворення активної ліпази в клітинах, підсилює процеси ліпогенезу, сприяє перетворенню вуглеводів у жири, пригнічує окиснювання вищих жирних кислот у тканинах (табл. 12.1.).
Таблица 12.1.
Вплив деяких факторів на мобілізацію жирних кислот із жирової тканини
| Фактор | Характер впливу | Передбачуваний механізм дії |
| Катехоламіни, глюкагон, тироксин, глюкокортикоїди | Посилення | Активація аденілатциклази |
| СТГ, АКТГ (гормони гіпофізу) | » | Посилення синтезу аденілатциклази і гормоночутливої ліпази |
| Стрес, фізичне навантаження, голодування, охолодження | » | Стимуляція секреції катехоламінів и пригнічення секреції інсуліну |
| Простагландини | Пригнічення | Ослаблена дія катехоламінів на аденілатциклазу, пригнічення аденілатциклази |
| Інсулін | » | Гальмування вивільнення жирних кислот в результаті активації гліколізу в жировій тканині; активація фосфодиестерази Ц-АМФ |
Статеві гормони також виражено діють на жировий обмін: при їхньому недоліку збільшується синтез і відбувається гальмування розпаду жирів, що є однією з причин збільшення маси тіла людей похилого віку, у яких відзначається вікове зниження функції статевих залоз.
Важливе значення в регуляції обміну ліпідів має співвідношення синтезу триацилгліцеролів і фосфоліпідів у тканинах, особливо в печінці. Це пояснюється тим, що жирні кислоти необхідні як для синтезу триацилгліцеролів, так і фосфоліпідів. Триацилгліцероли здатні відкладатися в резерв і при надлишковому накопиченні можуть викликати порушення діяльності кліток, аж до їхньої загибелі. Фосфоліпіди є гідрофільними сполуками: вони виводяться з печінки і доставляються до інших органів і тканин, що стимулює їх синтез і відповідно знижує утворення триацилгліцеролів.
У нормі вміст загальних ліпідів у крові складає 400-800 мг/л. Він змінюється в залежності від статі, віку, характеру і режиму харчування, рівня фізичної активності.
Порушення ліпідного обміну можуть наставати вже в процесі переварювання й усмоктування жирів унаслідок захворювань травного тракту. Крім того, вони можуть бути зв'язані з недостатнім надходженням у кишечник ліпази соку підшлункової залози або жовчі.
При спадкоємному захворюванні, обумовленому недостатньою активністю ліпопротеїнліпази крові, порушується перехід жирних кислот з хіломікронів плазми в "жирові депо". У плазмі збільшується вміст хіломікронів, внаслідок чого вона здобуває молочний колір. Підвищення рівня ліпідів у крові (гіперліпемія) може бути викликано фізіологічними причинами, наприклад, прийомом їжі (аліментарна гіперліпемія). Гіперліпемії виникають нерідко при цукровому діабеті, захворюваннях підшлункової залози (панкреатити), печінки (гепатити), нирок (нефрози). У їхній основі лежать порушення енергетичного обміну, зв'язані з недостатнім використанням вуглеводів і посиленим окиснюванням жирів. При цьому активізуються процеси мобілізації жиру (триацилгліцеролів) з "жирових депо". Він надходить у кров (транспортна гіперліпемія) і доставляється до тих органів, що мають недолік в енергії.
Гіперліпемії спостерігаються при деяких отруєннях, порушенні функції щитовидної, статевих і надниркових залоз.
У ряді випадків підвищується вміст ліпідів у сечі (ліпурія). Вона може виникати, наприклад, після їжі, особливо після прийому великої кількості риб'ячого жиру. Накопичення ліпідів у сечі відзначається при цукровому діабеті, туберкульозі легень, хворобах нирок, отруєннях, пухлинах підшлункової залози, інфекційних і гнійних процесах.
Гіполіпемія - зменшений вміст ліпідів у крові, спостерігається при цирозі печінки і зниженій функції щитовидної залози.
Недостатнє якісне і кількісне надходження ліпідів з їжею приводить до розвитку авітамінозів і гіповітамінозів жиророзчинних вітамінів. Так, зниження споживання з їжею рослинної олії приводить до появи ознак недостатності вітаміну F.
До захворювань, в основі яких лежать порушення обміну ліпідів, відносяться ожиріння, жирова дистрофія печінки, атеросклероз. У людей, що страждають на ожиріння, підсилюються процеси синтезу жирних кислот і триацилгліцеролів, що обумовлює їхнє накопичення і відкладення в клітинах. Жирова дистрофія печінки характеризується накопиченням надлишкової кількості триацилгліцеролів у її клітинах, що веде до дегенеративних змін клітин і порушенню їхніх функцій. У печінці активно відбувається обмін жирів, у тому числі два взаємозалежних процеси - синтез триацилгліцеролів і синтез фосфоліпідів. Посилення синтезу триацилгліцеролів гальмує утворення фосфоліпідів, і навпаки. При цьому триацилгліцероли легко відкладаються в печінкових клітках, а фосфоліпіди транспортуються до тканин, де беруть участь у різних процесах. При недоліку ліпотропних речовин, необхідних для синтезу фосфоліпідів, відбувається його гальмування, а жирні кислоти йдуть на синтез триацилгліцеролів, що накопичуються в клітках печінки, викликаючи її дистрофію.
Харчові речовини беруть участь у нормалізації порушень обміну ліпідів. Джерелами рухливих метильних груп, необхідних для синтезу ліпотропних речовин, є метіонін, вітаміни U, В15, холін, лецитин, бетаїн. На метилювання багатьох сполук впливають фолацин і вітамін В12.
В окисленні ліпідів, у тому числі холестеролу, до кінцевих продуктів беруть участь ніацин, триптофан, з якого частково утворюється ніацин при наявності вітаміну В6, В12, вітаміни С, Р, ліпоєва кислота, лецитин, холін.
Випаданню холестеролу в осад з рідких середовищ організму (зумовлюючому розвиток атеросклерозу, утворення жовчних каменів) перешкоджають поліненасичені жирні кислоти, що входять до складу рослинних олій. Важлива також їхня роль у зниженні формування ліпопротеїнів низкої щільності (які містять холестерол) і посиленні руйнування цих часток у крові. Вітамін С попереджає зменшення кількості ліпопротеїнів високої щільності, у складі яких холестерол швидше метаболізується, чим у частках низької щільності. Усмоктування цього ліпіду з кишечнику в кров гальмується b-ситостеролом, що міститься в рослинних жирах. Виведення холестеролу з організму через товсту кишку стимулюють вітаміни групи В (особливо тіамін), інозит, ПНЖК, а також магній, баластові речовини, сорбіт, ксиліт. Протисклеротичну дію мають деякі морепродукти внаслідок вмісту в них ПНЖК і йоду.
12.5. ПСУВАННЯ ЛІПІДІВ
Біохімічне (ферментне) прогіркання жирів відбувається під впливом ферментів плісеней: синтезована ними ліпаза викликає гідроліз жиру. Жирні кислоти, що утворюються при цьому, піддаються b-окисленню і наступному декарбоксилюванню.
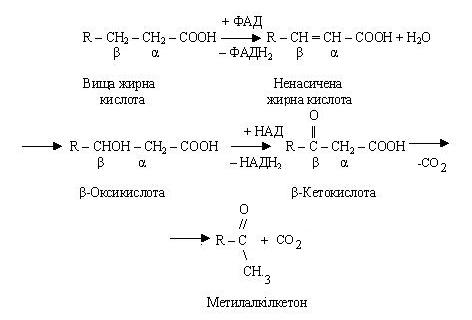
Кінцевий продукт цієї реакції - метилалкілкетон - додає жиру гіркий смак. Процес сповільнюється при низьких температурах і цілком припиняється при високих, у зв'язку з чим жири варто зберігати в холодильнику, а при необхідності тривалого збереження - перетоплювати.
Хімічне прогіркання (автоокислення) жирів відбувається при контакті з оксигеном повітря. Цей процес йде повільніше при низьких температурах, у середовищі інертних газів, у вакуумі і різко прискорюється при нагріванні (термоокислення).
Автоокислення - самокаталізуючийся процес, тому що нові радикали, маючи дуже високий рівень енергії, стимулюють процес окиснювання.
Первинними продуктами окиснювання є гідропероксиди. Вторинними - спирти, альдегіди, кетони, кислоти. Крім того, при автоокисленнні відбуваються зміни в положенні подвійних зв'язків у молекулах ненасичених жирних кислот (ізольована система подвійних зв'язків перетворюється в сполучену). Це прискорює окиснювання і полімеризацію жиру, особливо при нагріванні, і порушує структуру ессенціальних жирних кислот.
Прискорює автоокислення і дія світла, тому що ультрафіолетові промені постачають енергію, достатню для розщеплення зв'язку С-Н і для розкладання гідропероксидів з утворенням вільних радикалів.
Автоокислення активізується іонами важких металів (Pb, Cu, Fe, Cd) і сплавів (нержавіюча сталь), що можуть попадати в жир з матеріалів посуду й устаткування, особливо з місць, підданих корозії.
Температура вище 200 °С викликає сильне розкладання (піроліз) жирів з утворенням високотоксичних речовин, зокрема (при обвуглюванні) канцерогенних поліциклічних вуглеводнів: 3,4-бензпирену й інших (усього більш 200 летучих продуктів).
У нагрітих жирах виявлені продукти термоокислювальної деструкції, причому в оліях вміст летучих продуктів окиснювання вище, ніж у твердих жирах. У результаті цього процесу утворюються летучі карбонові кислоти (С2 - С18): моноєнові (C4 - C18), дієнові (С8 - С10), альдегідокислоти (С8 - С14), оксикислоти (С6 - С16), кетокислоти (С6 - С12), двохосновні кислоти (С6 - С11), а також алкани, алкадієни, спирти, альдегіди, кетони, складні ефіри, ароматичні і гетероароматичні сполуки. Крім того, у нагрітому жирі міститься акролеїн (акриловий альдегід, "кухонний газ"), що є продуктом термічної дегідратації гліцеролу.
Багато продуктів термоокислення жирів токсичні, викликають роздратування травного тракту людини. Найбільш виражений вплив, що ушкоджує, мають окислені жирні кислоти, що містять внаслідок окиснювання кисневмісні функціональні групи (ОН, С=О і ін.).
Ознакою полімеризації є збільшення в'язкості і молекулярної маси жиру при тривалому нагріванні.
До нелетучих продуктів термоокислення і термополімерізації відносяться циклічні жирні кислоти.
При оцінці якості нагрітих жирів використовують показник - фритюрну стійкість, тобто час (у годинах), протягом якого в жирі, що нагрівається, накопиться 1 % окислених жирних кислот. Жир вважають абсолютно зіпсованим, якщо в його масі хоча б 1 % складають окислені жирні кислоти. Цей показник залежить від ступеня контакту жиру з оксигеном, виду жиру, ступеня гідролізу і вмісту гідропероксидів у вихідному жирі.
Автоокислення ліпідів пригнічують сполуки, здатні швидко зв'язувати вільні радикали. Їх називають антиоксидантами (антиокислювачами) і застосовують для уповільнення окиснювання. Антиокисну активність має ряд природних сполук: фосфатиди, деякі вітаміни (Е, С, Р). Вони окисляються легше жирів, зв'язують вільні радикали і уповільнюють окиснювання жирів. Так, токофероли стабілізують різні жири, а також сухі молочні вершки. Антиокисні властивості властиві речовинам, виділюваним з маслянистої сировини, злаків, зрілих олив і їхнього соку. Вони зберігають свою активність і в продуктах переробки, тому антиокисну дію мають сирі соєва, бавовняна, пальмова, кунжутна олії, макуха льону, сої, арахісу, тонко розмелені обсмажені і знежирені боби какао, водні витяжки зі знежирених сої, арахісу, какао. Антиокислювачі виділені також з лушпайки бобів какао, насінь соняшника.
Антиокисну активність мають і меланоїдини, що утворюються в результаті взаємодії редукуючих цукрів з амінокислотами та іншими азотвмісними сполуками. Пряності в кількості 0,2 % маси лярду або жиру в майонезі затримують окиснювання жирів.
Поряд із природними антиокислювачами в харчовій промисловості застосовують і штучні. До них відносяться феноли і їхні похідні (крезоли, нафтоли, алкілпохідні фенолу й ін.). Найбільш розповсюдженими антиоксидантами цієї групи є бутилгідроокситолуол-бутилокситолуол (БОТ) і бутилгидроксианізол-бутилоксианізол (БОА), які використовуються для уповільнення окиснювання ліпідів у випечених виробах, кондитерській масі та ін.
Ефективними антиокислювачами виступають також госіпол - барвна речовина сирої бавовняної олії; 2,4,5-тригідрооксифеноли, ароматичні кислоти з двома-трьома гідроксилами в кільці (гідроксикавова, галова, 2,4,5-триоксибезойна, аміни, сірчисті сполуки, євгенол, кремнійорганічні сполуки).
При високій температурі (наприклад, при жарці) дія антиокислювачів знижується через їхню летючість і термічну нестійкість. Встановлено: чим нижче ступінь окиснювання вихідної сировини, тим вище його фритюрна стійкість. Отже, поза залежністю від того, чи буде жир використовуватися для жаркі, у нього обов'язково вводять антиоксиданти.
У харчовій промисловості антиокислювачі широко застосовують для стабілізації жирів і рослинних олій, жировмісних продуктів (ковбас, баликів), яловичого і сосискового фаршів, біфштексів. Додавання їх у фритюр не уповільнює окиснювання жирів у готових виробах.
Дії антиокислювачів сприяють синергісти - речовини, що підсилюють вплив антиокислювачів: лимонна і аскорбінова кислоти, деякі амінокислоти, аміни, фосфорна кислота, вуглеводи, сірковмісні сполуки.
З дубової кори, листів чаю, чорнильних горішків одержують галову кислоту, складні ефіри і солі, що відрізняються бактерицидними властивостями.
КОНТРОЛЬНІ ПИТАННЯ І ЗАВДАННЯ:
1. Що таке ліпіди і які їхні функції в організмі?
2. У складі свинячого жиру виявлені трипальмітин, триолеїн, пальмітодиолеїн. Напишіть їхні формули.
3. У кокосовій і пальмовій оліях знайдені стеародипальмітин, олеодипальмітин. Напишіть їхні формули.
4. У яєчному жовтку виявлені фосфоліпіди. Напишіть їхні формули і перелічите основні функції.
5. Який стероїд тваринного походження сприяє розвиткові атеросклерозу кровоносних судин?
6. Напишіть формулу холестеролу і вкажіть його значення для організму людини.
7. Назвіть основні функції ліпопротеїнів, гліколіпідів.
8. Які продукти є джерелами поліненасичених жирних кислот?
9. Від чого залежить характер і ступінь зміни жирів при технологічній обробці?
10. Присутність яких речовин каталізує процеси окислення жирів при технологічній обробці?
11. Як відбувається розщеплення ліпідів у шлунково-кишковому тракті людини?
12. Який склад жовчі і її роль у розщепленні ліпідів?
13. Яким перетворенням у клітці піддається гліцерол?
14. Напишіть реакції b-окиснювання вищих жирних кислот.
15. Який енергетичний ефект b-окиснювання вищих жирних кислот?
16. Які фактори здійснюють регуляцію ліпідного обміну?
17. Охарактеризуйте порушення обміну ліпідів.
Тести для контролю знань і самопідготовки
1. Які функції виконує холестерин в організмі людини?
| o | а) бере участь у побудові біологічних мембран |
| o | б) входить до складу цитоплазми тощо |
| o | в) із холестерину синтезуються жовчні кислоти, кортикостероїди, статеві гормони |
| o | г) усі зазначені функції |
2. Чи синтезується холестерин в організмі вищих тварин і людей?
| o | а) не синтезується |
| o | б) синтезується із бутирату |
| o | в) синтезується із КоА-ацетилу |
| o | г) синтезується із малонової кислоти |
3. Як задовольняються потреби організму в поліненасичених ЖК?
| o | а) за рахунок біосинтезу |
| o | б) за рахунок харчових ЖК |
| o | в) за рахунок біосинтезу і харчових ЖК |
| o | г) організм не потребує поліненасичених ЖК |
4. Як включаються в катаболізм нейтральні жири?
| o | а) розщеплюються ліпазою на гліцерин і ЖК |
| o | б) включаються молекулою триацилгліцериду |
| o | в) поступово відщеплюють ЖК від триацилглідериду |
| o | г) включаються тільки ЖК |
5. Як окислюється гліцерин?
| o | а) через гліцеринову кислоту |
| o | б) через гліцераль-3-фосфат |
| o | в) через діоксиацетон-3-фосфат |
| o | г) не окислюється |
6. Як активуються ЖК, вступаючи в катаболізм?
| o | а) фосфорилюються за рахунок АТФ |
| o | б) не змінюють структури |
| o | в) утворюють ацил-КоА за рахунок енергії АТФ |
| o | г) конденсуються з КоА без затрат енергії |
7. Хто вперше розкрив механізми b-окислення?
| o | а) Г. Кребс |
| o | б) В. Палладій |
| o | в) Е. Фішер |
| o | г) Ф. Кнооп |
8. У чому полягає β-окислення?
| o | а) у повторенні прямого і непрямого дегідрування, яке завершується утворенням ацетилу-КоА |
| o | б) у повторенні дегідрування і гідратації |
| o | в) у повторенні гідратації і дегідрування |
| o | г) у повторенні дегідрування, гідратації та декарбоксилювання |
9. Як ацетил-КоА включається в енергетичний обмін?
| o | а) включається в циклі трикарбонових кислот |
| o | б) включається в гліколіз |
| o | в) самостійно окислюється до СО2 + Н2О |
| o | г) перетворюється на піровиноградну кислоту |
10. Який сумарний вихід АТФ під час перетворення ацетил-КоА до СО2 і Н20 в циклі трикарбонових кислот?
| o | а) 131 |
| o | б) 12 |
| o | в) 123 |
| o | г) 132 |
11. Яка речовина утворюється в одному циклі β-окислення?
| o | а) СО2 + Н20 |
| o | б) молочна кислота |
| o | в) ацетил-КоА |
| o | г) піровиноградна кислота |
12. Скільки ацетил-КоА утворюється внаслідок повного β-окислення стеаринової кислоти?
| o | а) 8 |
| o | б) 6 |
| o | в) 9 |
| o | г) 12 |
13. Які патологічні стани та хвороби супроводжуються метаболічним кетозом?
| o | а) голодування - повне або вуглеводне (нестача ЩУК) |
| o | б) цукровий діабет (порушення вуглеводного обміну, нестача ЩУК) |
| o | в) зміни дієти (порушення метаболізму - нестача ЩУК) |
| o | г) усі зазначені стани та хвороби |
14. Чому у разі нестачі ЩУК нагромаджуються ацетонові тіла?
| o | а) блокується окислення ацетилу-КоА |
| o | б) ацетил-КоА конденсується у ацетооцтову кислоту |
| o | в) припиняється перетворення ацетооцтової та β-гідроксимасляної кислот |
| o | г) за рахунок усіх зазначений процесів |
15. Як утворюється ацетон?
| o | а) у процесі β-окислення |
| o | б) під час декарбоксилювання ацетооцтової кислоти |
| o | в) під час конденсації 2 КоА-ацетилів |
| o | г) внаслідокa-окислення |
16. Як запобігти утворенню ацетонових тіл?
| o | а) уникати порушень вуглеводного обміну |
| o | б) уникати порушень β-окислення |
| o | в) уникати перевантаження організму жирами |
| o | г) дотримуватися у харчуванні певної пропорції: вуглеводи/жири |
17. Які шляхи боротьби з ацетонемією?
| o | а) відновлення порушеного вуглеводного обміну |
| o | б) введення глюкози хворому на цукровий діабет |
| o | в) введення інсуліну людині, що голодує |
| o | г) виведення із раціону жирів |
18. Які органічні сполуки їжі сприяють накопиченню ацетонових тіл у хворого на цукровий діабет?
| o | а) жири |
| o | б) вуглеводи |
| o | в) білки (кетогенні АК) |
| o | г) усі зазначені сполуки |
19. Яким чином відбувається регуляція ліпідного обміну?
| o | а) через вегетативну нервову і ендокринну системи |
| o | б) шляхом проміжного обміну |
| o | в) утворенням кінцевих речовин обміну |
| o | г) нервова система не бере участі у регуляції ліпідного обміну |
20. Які порушення ліпідного обміну викликає негативний емоційний стрес?
| o | а) порушення ліпідного обміну не пов'язані з емоційним стресом |
| o | б) збільшення виходу катехоламінів - мобілізація ліпідів - схуднення |
| o | в) ожиріння |
| o | г) ацетонемія |
21. Яка причина виникнення кетозу?
| o | а) нездатність ацетил-КоА включатися у синтез ЖК |
| o | б) нездатність ацетил-КоА включатися у синтез холестерину |
| o | в) нездатність ацетил-КоА вступати в ЦТК |
| o | г) усі зазначені причини |
РОЗДІЛ 13. БІОХІМІЧНІ ТА ІНШІ ЗМІНИ, ЩО ВІДБУВАЮТЬСЯ З ОСНОВНИМИ РЕЧОВИНАМИ В ПРОЦЕСІ ЗБЕРІГАННЯ І ТЕХНОЛОГІЧНОЇ ОБРОБКИ ХАРЧОВОЇ ПРОДУКЦІЇ
13.1.ЗМІНА СТАНУ ТА ВМІСТУ ВОДИ
Як відомо, вода – найбільш розповсюджена сполука в живих організмах. Вона – реакційно здатна сполука, вона дуже сильно відрізняється від більшості інших рідин складом і властивостями. Вода і продукти її дисоціації визначають структуру і біологічні властивості білків, нуклеїнових кислот, ліпідів та інших органічних речовин.
Вода у рідкому стані уявляє собою полімер, будова якого зумовлює специфічні фізичні властивості:
- висока температура плавлення, кипіння, випару;
- великий поверхневий натяг.
- при нагріванні від 0 до 40С її об’єм зменшується, максимальна густина досягається при 3,980С;
- при замерзанні об’єм води збільшується. Так об’єм льоду більше об’єму води, із якої він утворився; густина льоду 0,92 г/см3, він легше води;
- температура замерзання води знижується при збільшенні тиску;
- відрізняється високою діелектричною сталою і одним із самих великих поверхневих натягів;
- відрізняється високим ступенем змочування;
- здатна підніматися високо вгору в тонких капілярах і прилипати до поверхонь багатьох тіл;
- є універсальним розчинником багатьох речовин;
- володіє надмірно великою питомою теплоємністю.
Зберігання. Вода входить до складу всіх харчових продуктів. При виробництві харчових продуктів необхідно враховувати вміст в продуктах води, її фізіологічну роль в клітинах, вплив на зберігання продукту, характер зв’язку з матеріалом, а також мати уяву про формування кристалів при заморожуванні. У тваринних і рослинних тканинах вода є найбільш рухливим компонентом. Так, вміст води у свіжому оселедці в залежності від віку, статі, району і часу лову коливається в межах від 51,0 до 78,3 %, у трескових риб – від 70,6 до 86,2 %. У картоплі в залежності від сорту, району вирощування, грунту і вегетаційного періоду кількість води складає 67 - 83 %.
Велика частина води у свіжих овочах і плодах перебуває у вільній, рухливій формі, незначна частина (до 5 %) у зв’язаному стані. При випаровуванні води послаблюється тургор клітин і відбувається зів’янення тканин, посилюються процеси розпаду, збільшується витрата на дихання, порушується енергетичний баланс, стійкість овочів і плодів до ураження мікроорганізмами знижується.
Основна причина зменшення маси рослинної сировини – випаровування води. У середньому 2/3 втрат маси овочів і плодів при зберіганні відбувається за рахунок випаровування води і 1/3 за рахунок втрати органічни речовин (при температурі до 50С), при більш високих температурах ці втрати збільшуються. Так, втрати маси бульб картоплі при температурі 00С за рахунок випаровування води втричі більші, ніж у результаті дихання, а при 130С – у 1,3 рази.
У більшій частині харчової сировини (м’ясо, субпродукти, птиця, риба та ін.) вміст води і розчинних речовин змінюється на наступних етапах обробки:
- зберігання;
- при розморожуванні сировини і зберігання напівфабрикатів із неї;
- в процесі вимочування солоної продукції.
Функції організму тварин і рослин здійснюються тільки при достатньому вмісті в тканинах води, так плоди, овочі при втраті води в кількості 5-7 % в’януть і втрачають свіжість. Специфіка хімічного складу овочів і плодів зумовлює необхідність їх зберігання при певних умовах, наведених в таблиці 13.1.
Таблиця 13.1
Умови і режими зберігання овочів і плодів
| Вид продукту | Умови і режими зберігання | ||||
| Умови | Характер освітлення | Температура, 0С | Відносна вологість, % | Термін зберігання | |
| Капуста білокачанна | Контейнери, стелажі, ящики | Розсіяне | 0-10С | 90-97 | 4-7 міс |
| Капуста квашена | Деревяні або бетонні чани – домники. Вона повинна бути вкрита розсолом | Затемнене | 0-10С | 90-95 | 4-7 міс |
| Морква | Контейнери, ящики | Затемнене | 0-10С | 90-97 | 3-7 міс |
| Листковий і качанний салат, цибуля-порій, зелень петрушки, сельдерея, укропу | Лотки, ящики | Розсіяне | 00С | 95-97 | Від 1 тижня (зелень) до 3 міс |
| Томати | Лотки, ящики | розсіяне | 85-90 | Від 1 тижня до 1 міс | |
| Огірки | Лотки, ящики | затемнене | 10-12 | 90-95 | До 1 міс |
Продовження таблиці 13.1
| Перець солодкий, баклажани | Лотки, ящики | розсіяне | 0-1 | 80-85 | До 20 днів |
| Яблука | ящики | затемнене | 0-1 | 90-95 | До 1 міс літні сорти, до 10 міс зимні сорти |
| Цитрусові | ящики | затемнене | 1-5 | 85-90 | Від 1 міс до 3 міс |
| Ягоди, виноград | Лотки, ящики | затемнене | 0-2 | 85-95 | Від 1 дня до 2 тижнів |
Вода необхідна для хімічних, біохімічних реакцій і колоїдних процесів, що протікають у тваринних і рослинних тканинах під час їхньої переробки. Втрата води тваринами в межах 15-20 % призводить до їхньої загибелі.
Окрім наявності в харчових продуктах (м’ясо, риба, молоко, плоди, овочі) значної кількості води 70-93 %, вміст вуглеводів, азотистих речовин і жирів створює умови для розвитку мікроорганізмів, тому така продукція швидко псується. Між тим борошно, крохмаль, крупи, сушені овочі, плоди і ягоди зберігають при певних умовах на протязі тривалого часу.
Багато продуктів, насамперед, тваринного походження з метою збільшення терміну зберігання підлягають заморожуванню. В залежності від температури охолодження заморожених продуктів більша або менша частина води в них знаходиться у вигляді льоду. Волога, що утворюється в результаті відтаювання льоду, при розморожуванні продуктів може залишатися в них або виділятися в оточуюче середовище, що залежить від фізико-хімічних особливостей продуктів.
При заморожуванні м’язової тканини тварин і риб кристали льоду утворюються переважно в тканинній рідині між м’язовими волокнами і в меншому ступені в волокнах. Чим повільніше йде процес заморожування, тим в більшому ступені замерзає тканинна рідина між м’язовими волокнами і тим значніше зневоднюються м’язові волокна.
При відтаюванні м’язова тканина наземних тварин і риби веде себе по-різному.
Для м’яса в тому чи іншому ступені характерне відновлення структури після відтаювання, тобто поглинання вологи з тканинної рідини м’язовими волокнами. На відновлення структури м’яса впливають:
- швидкість заморожування;
- тривалість зберігання;
- тривалість і умови розморожування.
Правильно проведене розморожування дозволяє отримати м’ясо близьке за якістю до остиглого або охолодженого.
Найбільш сприятливими умовами і терміном розморожування вважаються для четвертин яловичини від 3 до 5 діб, для свинячих і баранячих туш – від 2 до 3 діб при температурі від 0 до 50С.
Для субпродуктів швидкість відтаювання не має значення, виключенням є серце, яке для попередження значних втрат м’ясного соку відтаюють повільно. Печінка, клітинні оболонки якої при заморожуванні руйнуються, незалежно від термінів розморожування втрачає близько 11-12 % соку.
У риби м’язова тканина не відновлює своєї структури при розморожуванні, тому відтаювання її (крім риб осетрових порід і філе) проводять швидко у воді і при температурі не вище 250С. При цьому риба поглинає 5-10 % вологи і втрачає близько 0,25 % органічних і 0,1 % мінеральних речовин. Втрати філе риби при розморожуванні на повітрі складають 4-7 %.
Напівфабрикати із риби і м’яса. Напівфабрикати із риби і м’яса, що підлягають швидкому відтаюванню, окрім втрат, пов’язаних з первинною обробкою, втрачають м’ясний сік при зберіганні. Порційні шматки вагою 100-140 г, укладені в один ряд за 6 годин при зберіганні при 18-200С втрачають близько 1 % м’ясного соку, а великошматкові напівфабрикати із м’яса, які після прискореного відтаювання добу витримувалися при температурі 2-40С за 24 год зберігання в холодильнику втрачають до 0,6 % соку. М’язова тканина риби слабкіше звязує воду, тому для попередження значних втрат сока її фіксують.
Вимочування. Солонина містить 6-12 %, а середньосолона риба 10-14 % кухонної сілі. Для видалення надлишку солі солоні продукти вимочують у воді, яку періодично міняють. Залишкова кількість солі повинна бути в сировині не більше 3-4 %). Вага продукції при цьому збільшується, але разом із кухонною сіллю у воду переходить значна частина розчинних поживних речовин: білків, екстрактивних та мінеральних речовин, тому за смаковими якостями і поживною цінністю ці продукти поступаються свіжим.
Як вже було сказано, вміст води в харчових продуктах відіграє важливу роль при формуванні і збереженні їхньої якості. Так, вода впливає на консистенцію продуктів (рідка, напіврідка або тверда), на стан поверхні і внутрішню будову ряду продуктів.
На збереження харчових продуктів впливає не тільки загальна кількість води, але і співвідношення вільної і зв’язаної, яке характeризує активність води.
Активність води виражається відношенням тиску водяних парів над продуктом до їхнього тиску над поверхнею чистої води при одній і тій же температурі.
Цей показник означає доступність води для фізичних, хімічних, фізико-хімічних і мікробіологічних процесів. Від величини активності води залежать:
- терміни зберігання м’яса і м’ясопродуктів;
- стабільність м’ясних консервів;
- формування кольору та запаху;
- втрати в процесах термообробки і зберіганні.
Цей показник дозволяє встановити взаємозв’язок між станом слабозв’язаної вологи в продукті і можливістю розвитку в ньому мікрорганізмів.
Між зв’язаною і вільною водою продуктів не спостерігається різкої межі. У рослинних і тваринних тканинах переважає вільна вода. Так, у м’язах тварин і риб основна частина води зв’’язана з гідрофільними білками за рахунок осмотичних (45-55 %), капілярних (40-45 %) сил, води змочування (0,8-2,5 %), а на частку зв’язаної води приходиться тільки 6,5-7,5 %. У плодах і овочах знаходиться до 95 % вільної води. Тому ці продукти сушать до вмісту залишкової вологості 8-20 %, тому що вільна вода з них легко видаляється.
При додаванні розчинних речовин, таких, як сіль, цукор, білкові інгредієнти, посилюється зв’язок вологи в харчовому продукті.
При видаленні вологи висушуванням, перетворенням вологи в лід в процесі заморожування також зменшується кількість слабозв’язаної води. Підвищення осмотичного тиску знижує парціальний тиск водяної пари і активність води зменшується. На цих ефектах грунтуються традиційні способи консервування продуктів.
Активність води можна змінювати, підбираючи сировину і рецептури з урахуванням використаної кількості кухонної сілі і жиру. Створення оптимальних умов зневоднення ковбас в процесі дозріванні також дає можливість регулювати активність води. В дозрілих ковбасах ріст небажаних мікроорганізмів стримується низькою активністю води, анаеробністю середовища, низьким значенням рН, наявністю хлориду і нітриту натрію, аттакож молочнокислої мікрофлори.
Ступінь впливу на активність води харчових добавок зменшується в наступному порядку: кухонна сіль, поліфосфат, цитрат, тартрат, гліцерин, лактоза, молочний білок, жир. Добавки, молекули яких мають більший ступінь дисоціації, сильніше знижують активність води, ніж макромолекулярні речовини.
Форми зв’язку води з речовинами і структурними елементами харчового продукту:
- фізико-механічна (волога змочування, волога в макро- і мікрокапілярах);
- фізико-хімічна (волога, набрякання, адсорбційна);
- хімічна (іонна і молекулярна).
Переважають перші дві форми зв’язку, хімічний зв’язок у продуктах зустрічається рідко.
Фізико-механічний зв’язок обумовлений утриманням вологи в проміжках структури (імобілізаційна вода), у мікро- і макрокапілярах і прилипанням її до поверхні часток або продукту (змочування), вилучається з матеріалу випарюванням або механічним способом (віджим, центрифугування та ін.), при цьому основна маса води знаходиться у вільному стані і не змінює своїх властивостей.
Волога змочування – волога у вигляді дрібних крапель утримується силами поверхневого натягу на поверхні розрізу тканин продуктів. Вода досить легко видається з продукту, тому що слабозв’язана із субстратом.
Капілярна волога заповнює пори і капіляри (наприклад, м’яса, фаршу). Її кількість залежить від ступеня капілярності сировини, матеріалу. При нарізанні м’яса, риби, плодів, овочів може відбуватися часткова втрата м’язового, плодового та овочевого соку, що має високу харчову цінність. В м’ясі роль капілярів виконують кровоносні і лімфатичні судини. Капілярна волога впливає на об’єм і соковитість продукту; чим вище капілярний тиск, тим міцніше капілярна волога, пов’язана з матеріалом. Капілярний тиск, в свою чергу, визначається розміром капілярів; чим менше діаметр капіляра (мікрокапіляр), тим він вище і тим міцніше утримується вода.
Навіть при однаковій формі зв’ язку вологи її міцність і вплив на властивості тканин неоднакові.
В технологічній практиці вологу за формою її звязку з м’ясом спрощено ділять на міцно зв’язану, слабо зв’язану корисну і слабко зв’язану надлишкову.
Міцно зв’язана волога – це в основному адсорбційна, а також волога мікрокапілярів і частина осмотичної вологи.
Слабко зв’язана корисна волога пом’якшує (пластифікує продукт), cтворюючи сприятливу консистенцію і сприяє засвоєнню їжі.
Слабко звязана надлишкова волога може відділятися в процесі технологічної обробки у вигляді бульону.
При виготовленні ковбас міцно зв’язана волога повинна складати приблизно 1/3 всієї рідини. У випадку виготовлення ковбаси із замороженого м’яса, яке довго зберігалося, частина вологи виявляється у вигляді слабозв’язаної надлишкової і консистенція продукту гірша, відбувається відділення бульйону і зменшується вихід продукту. При більшій кількості міцнозв’язаної вологи (більше 1/3 частини) продукт отримують дуже щільним, при цьому зменшується випаровування вологи. Так, при обжарці ковбас втрати за рахунок випарення вологи складають 7-8 %. При сушінні ковбасних виробів (особливо сирокопчених, сировялених) бажано, щоб міцнозв’язаної вологи було менше. Впливати на кількість вологи того чи іншого зв’язку можна змінюючи рН.
Водозвязуюча здатність сировини (м’ясо, риба та ін.) визначає її властивості на різних стадіях технологічної обробки і впливає на водозв’язуючу здатність готових продуктів, їх якість і вихід. Оскільки переважними компонентами (м’яса, риби) є м’язова і сполучна тканини, то їх водозв’язуюча здатність має найбільше значення.
Водозв’язуюча здатність м’яса залежить в основному від стану білків; жири лише в незначній мірі утримують вологу. Основна частина води (близько 90 %) міститься у волокнах м’язової тканини, при цьому в міофібрилах її більше, в саркоплазмі менше, тому водозв’язуюча здатність м’язової тканини в першу чергу визначається властивостями і станом білків міофібрил (актин, міозин, актоміозин). В сполучній тканині води менше, вона пов’язана головним чином з колагеном.
Окрім капілярного звязку існує адсорбційна і осмотична форми.
Фізико-хімічний зв’язок зумовлений вологою в гідратних оболонках або осмотичним утримуванням в клітинах у несуворо визначених співвідношеннях; вилучається з матеріалу випарюванням, десорбцією (адсорбційна), або внаслідок різниці концентрацій (осмотична).
Адсорбційна волога – найбільш міцно зв’язана волога, яка утримується за рахунок сил адсорбції, головним чином, білками. Диполі води фіксуються гідрофільними центрами білків. Число заряджених груп білку залежить від ряду факторів і може зменшуватися навіть до нуля (ізоелектрична точка).
Водозвязуюча здатність білків тим вища, чим більше інтервал між рН середовища і ізоелектричною точкою. Так, якщо тварина перед забоєм отримала стрес, то автолітичні і гліколітичні процеси в м’ясі тварин посилюються, а рН різко зсувається в кислу сторону, тобто наближається до ізоелектричної точки. Таке мясо втрачає багато соку, а також має понижєену гідратацію. Туша тварини стає особливо водянистою при рН 5,2-5,5. Число неіонізованих полярних груп звичайно залишається незмінним, зберігається здатність білка зв’язувати деяку кількість води і в ізоелектричній точці.
Число груп, які фіксують вологу за рахунок сил адсорбції залежить від взаємодії білків. Наприклад, в процесі розвитку післязабійного задубіння це пов’язано з утворенням актоміозину із актину і міозину.
Осмотична волога утримується в незруйнованих клітинах за рахунок різниці осмотичного тиску з обох боків клітинних оболонок (напівпроникних мембран) і в клітинних мембранах. В міжклітинних просторах, як і в тканинах з неклітинною структурою, роль напівпроникної перетинки виконує каркас білкових гелів, у вічках якого міститься вода.
Осмотична волога міститься в м’ясі, рибі тим більше, чим менше руйнування напівпроникних мембран або структурних утворень, які виконують їх роль. Вона частково виходить із м’яса при зануренні його в розчин з більш високим осмотичним тиском (посол) і при тепловій денатурації білків. Кількість осмотичної вологи впливає на пружність тканин.
Теплова обробка. В продуктах тваринного і рослинного походження при всіх видах теплової обробки відбувається зміна вмісту води. Величина втрат залежить від хімічного складу сировини і способу обробки.
Поглинання води колагеном лише частково компенсує втрату її мязовими білками. Тому мясо – яловичина, свинина, - птиця, риба при тепловій обробці в більшому або меншому ступені зневоднюються. В рослинних продуктах зміни вмісту води залежить від кількості і стану білків, вуглеводів (насамперед крохмали) і способу термічної обробки. Так, при варінні картоплі втрати води складають від 3 до 5 %, а при жарінні до 50-60 %.
Процес виділення води з м’яса і риби протікає по-різному. Чим вище температура нагрівання м’яса, тим більше воно втрачає води. При нагріванні риби такої закономірності не спостерігають і максимальна кількість води виділяється при t 65-750С, з підвищенням температури втрати води зменшуються. Різниця вказує на те, що поглинання води колагеном компенсує втрати її мязовими білками риби в більшому ступені, ніж мяса.
Як при варінні, так і при повному прожарюванні різні види мяса втрачають близько 50 %, а різні види риби – близько 25 % води, що в них міститься. Але в характері виділення води є значна різниця. Під час варіння у воді вся волога, що міститься у тваринному продукті, надходить в оточуюче середовище в рідкому стані. При жарінні тільки частина вологи виділяється в рідкому стані, утворюючи сік. Основна її маса випарюється спочатку на поверхні шматка і в шарах, що безпосередньо прилягають до неї, а потім по мірі пргрівання в більш глибоких шарах. При варінні продуктів тваринного походження на пару, припусканні і тушінні виділення вологи відбувається менше, ніж при варінні у воді, але більше, ніж при жарінні.
Таким чином, вода в харчових продутах при переробці і зберіганні може переходити з вільної в зв’язану і навпаки, що викликає зміну властивостей сировини і готової продукції. Наприклад, при випіканні хліба, варінні картоплі, виробництві мармеладу, пастили, драглів, желе, кремів відбувається перетворення частини вільної вологи в адсорбційно зв’язану з колоїдними частками білків, крохмалю і інших речовин, а також зростає кількість осмотично утримуваної води. У соках із плодів, ягід, овочів змінюються форми зв’язку води в порівнянні з вихідною сировиною. При відмоканні мармеладу в результаті старіння драглів, при відтаванні замороженого м’яса і картоплі спостерігається перехід частини зв’язаної води у вільну.
13.2. ЗМІНИ ВМІСТУ МІНЕРАЛЬНИХ РЕЧОВИН
Мінеральні речовини, які містяться в харчових продуктах, при зберіганні і технологічній обробці змінюються мало, але значна їх частина може втрачатися, переходячи у воду при промиванні, замочуванні і варінні продуктів, або вилучатися разом із соком, який виділяється при жарінні м‘яса, птиці, риби.
Зберігання і механічна обробка. Вміст мінеральних речовин, які концентруються в основному в овочевій продукції в корі, у верхній частині клубнів у картоплі в процесі механічної обробки (очищення, зберігання) зменшуються.
Дані про загальну кілкьість сухих і мінеральних речовин, які переходять у воду при зберіганні в ній очищеної картоплі, наведені в таблиці 13.2.
Таблиця 13.2.
Втрати сухих і мінеральних речовин, очищеної картоплі при зберіганні у воді, % від маси картоплі
| Тривалість зберігання, год | Загальна кількість сухих речовин | Мінеральні речовини | ||
| Цілі клубні | Нарізані клубні | Цілі клубні | Нарізані клубні | |
| 0,07-0,16 | 0,47-0,49 | 0,01-0,02 | 0,04-0,05 | |
| 0,13-0,18 | 0,50-0,61 | 0,02-0,03 | 0,05 | |
| 0,15-0,23 | 0,62-0,72 | 0,03-0,04 | 0,08-0,09 |
При замочуванні бобових втрати мінеральних речовин складають 4-11 %. Втрати мікроелементів при замочуванні гороху в холодній воді складають 3,8-5,5 %, а при замочуванні в гарячій воді від 6 до 11 %. При замочуванні квасолі втрати цих речовин незначні. При бланшируванні, консервуванні і варінні зеленого гороху вміст в ньому магнію, калію, фосфору зменшується, а вміст кальцію збільшується (останнє зв‘язують з поглинанням його із води). Замочування бобових скорочує час варіння, в результаті чого загальні втрати не збільшуються. Втрати соку при відтаюванні м‘ясної туші можуть досягати до 10 %, в загальному об‘ємі втрати мінеральних речовин складають 1,1 %.
Денатураційні зміни призводять до зниження кількості мікроелементів, повязаних з білком. При цьому збільшується здатність полісахаридів квасолі і гороху звязувати окремі мікроелемнети. Так, після доведення квасолі до готовності вміст у вуглеводному комплексі марганця зростав з 22,8 до 44,4 %, міді – з 24,4 до 32,7, молібдену – з 15,8 до 26,1 %.
Теплова обробка. При варінні овочів руйнується шкіряний шар протоплазми. Розчинні в клітинному соку речовини отримують можливість вільно дифундувати в оточуюче середовище. Втрати їх можуть бути дуже великі. Так, при варінні очищеної картоплі у відвар переходить більш 20 % мінеральних речовин.
Значно знижує втрати мінеральних речовин варіння овочів в шкірці, жаріння і варіння паром і в мікрохвильових печах, пароконвектоматах. Дещо більше вилучається їх при припусканні і більше при варінні продуктів з повним зануренням в рідину. Щоб зменшити втрати мінеральних речовин при кулінарній обробці овочів, слід занурювати їх при варінні в киплячу воду, використовувати овочеві і круп‘яні відвари, брати рідини не більше, ніж це вимагається, щоб вона повністю вкривала продукти.
Втрати мікроелементів при варінні бобових коливають від 5 до 41 %, представлені в таблиці 13.3.
Таблиця 13.3
Втрати мікроелементів при варінні бобових
| Умови варіння бобових | Втрати в % до вихідного вмісту | ||
| Марганець | Мідь | Молібден | |
| Горох Варіння після замочування в холодній воді на протязі 8-10 год Варка після замочування в гарячій воді (900С) на протязі 2,5-3, год | 11,8 10,6 | 17,0 16,2 | 41,3 36,2 |
| Квасоля Варіння після замочування в холодній воді на протязі 8-10 год | 6,0 | 19,5 | 40,8 |
| Варіння після замочування в гарячій воді (900С) на протязі 2,5-3 год | 5,0 | 16,5 | 40,0 |
Кількість речовин, що виділяються із м‘яса в процесі варіння, підлягає значним коливанням. Однією із причин цього є відмінність у властивостях м‘яса, зумовлена породою, статтю, віком і ступенем вгодованості тварини, від якої отримане м‘ясо, має значення і термічний стан м‘яса. При інших рівних умовах остигле, охолоджене і морожене м‘ясо виділяє під час варіння неоднакову кількість розчинних речовин.
В багатьох випадках втрати розчинних речовин залежать від способу теплової обробки і від концентрації рідини. За даними Лобанова Д.І. виділення розчинних речовин при варінні м‘яса з різною кількістю води складає:
| Співвідношення кількості м‘яса і води | Загальна кількість розчинних речовин | в т.ч. мінеральні речовини |
| Без води | 1,99 | 0,57 |
| 1:1 | 2,34 | 0,63 |
| 1:2 | 2,57 | 0,75 |
| 1:3 | 2,87 | 0,87 |
| 1:5 | 2,88 | 0,90 |
При варінні яловичини виділяється в середньому (в % від маси м‘яса) до 0,55 мінеральних речовин. Згідно літературних даних, мінеральний склад концентрованих бульйонів із грудинки і стегнової частини туші яловичини в % від маси м‘яса відповідно складають 0,31 і 0,45, а в процентах від загальної кількості сухих речовин бульйону – 18 і 24 відповідно.
При варінні птиці в бульйон переходить 0,25 % мінеральних речовин від маси м‘яса.
Не нашли, что искали? Воспользуйтесь поиском: