ТОР 5 статей:
Методические подходы к анализу финансового состояния предприятия
Проблема периодизации русской литературы ХХ века. Краткая характеристика второй половины ХХ века
Характеристика шлифовальных кругов и ее маркировка
Служебные части речи. Предлог. Союз. Частицы
КАТЕГОРИИ:
- Археология
- Архитектура
- Астрономия
- Аудит
- Биология
- Ботаника
- Бухгалтерский учёт
- Войное дело
- Генетика
- География
- Геология
- Дизайн
- Искусство
- История
- Кино
- Кулинария
- Культура
- Литература
- Математика
- Медицина
- Металлургия
- Мифология
- Музыка
- Психология
- Религия
- Спорт
- Строительство
- Техника
- Транспорт
- Туризм
- Усадьба
- Физика
- Фотография
- Химия
- Экология
- Электричество
- Электроника
- Энергетика
МИТОХОНДРИЯЛАР құрылысы қызметі
СӘТТІЛІК
1.Клетка ілімі тарихының қалыптасуы Клетка (жасуша) – біздің планетамыздағы тірі ағзалардың құрылымы мен дамуының негізін құраушы бірлігі. Көп уақыттар бойы биология жануарлар мен өсімдіктердің құрылыстарының қасиеттерін, олардың көзге көрінетін макроскопиялық құрылысының негізінде зерттеп келді. Ағзалардың жасушалық құрылысын ашқаннан соң, клетканы тірі ағзалардың құрылымдық және функциональді бірлігі ретінде қарастырғаннан бастап, биология ағзалардың құрылысы мен қызметін неғұрлым терең зерттей бастады. Клеткалар жай көзге көрінбейді, сондықтан тірі ағзалардың клеткалық құрылымын оқып, зерттеу жұмыстары оптикалық аспаптардың жасалуы және жетілуімен тығыз байланысты. XVI ғ. аяғы мен XVIIбасында жаратылыстану ғылымдары саласында оптикалық аспаппен тәжірибе жүргізу қауырт дамыды. Кейбір деректерде алғашқы микроскопты голландиялық оптиктер Янсендер 1590 жылы жасап шығарған делінді 1609-1610 жж. Галилео Галилей ең алғаш оптикалық аспапты ойлап құрастырды, ол тек 1624 ж. ғана сол аспапты тәжірибелерде пайдалана алу деңгейіне жеткізді. Бұл аспап 35-40 ретке ұлғайтатын болды. Бір жылдан кейін И. Фабер осы аспапқа “микроскоп” атын берді. XVII ғ. 70 ж. Марчелло Мальпиги өсімдіктердің кейбір мүшелерінің микроскопиялық құрылысын зерттеді. 1682 ж. Н. Грю “Өсімдіктер анатомиясының бастамасы”деген еңбегін жазды.
XVII ғ. III жартысында голландиялық ғалым Антон ван Левенгуктың еңбектерінде жануарлар клеткасының құрылысы туралы зерттеулер орын алды. Ол микроскоп құрылысын жетілдіре отырып, жануарлар клеткаларын зерттеп, ұлпалар мен мүшелердің құрылысын қарастырды. 1696 ж. оның “Жетілдірілген микроскоптар көмегімен ашылған табиғат құпиялары” атты еңбегі жарық көрді. Левенгук алғаш рет эритроциттерді, сперматозоидтарды зерттеп, сипаттама берді, көзге көрінбейтін құпия әлем – микроағзаларды ашып, оларды инфузориялар деп атады. Левенгук ғылыми микроскопияның негізін қалаушы болып саналады. 1715 ж. Х.Г. Гертель микроскоп объектілеріне жарық түсіруге алғаш рет айнаны пайдаланады, дегенмен тек 1,5 ғасырдан кейін ғана Э. Аббе микроскоп үшін жарық түсіргіш линзалар жүйесін құрастырды. 1781 ж. Ф. Фонтана жануарлар клеткаларының небір құпияларын ашып, жануар клеткасын ядросымен қосып суретін салды. XIX ғ. I жартысында чех ғалымы Ян Пуркинье микроскоп техникасын жетілдіріп, клетка ядросын (“ұрық көбігі”) сипаттап, жануарлар мүшелерінің әр түрлі клеткаларын зерттеді. Ол клетка ішіндегі сұйықтыққа “протоплазма” деген ұғымды қолданды. 1831 ж. Роберт Браун жасуша ядросын маңызды да тұрақты құрылым ретінде сипаттап, “nucleus” – ядро ұғымын енгізді. 1838 ж. неміс ботанигі Матиас Шлейден цитогенез, яғни клетканың түзілуі теориясын енгізді.
Шлейден ағзадағы клеткалардың пайда болуы сұрағының басын ашты, сондай-ақ ол ядро – барлық өсімдік клеткаларының міндетті компоненті деген қорытындыға келді. Шлейденнің отандасы зоолог Теодор Шванн өсімдіктер және жануарлар ағзаларының клеткаларын салыстырып, олардың құрылысы ұқсас деп қорытындылады. Клетка туралы өздеріне дейінгі мәліметтерді жинақтап және өз зерттеулерінің нәтижелерін пайдалана отырып, ботаник М. Шлейден мен зоолог Т. Шванн жасуша теориясының негізін қалады. Клетка теориясының қағидалары Шлейденнің 1838 ж. “Өсімдіктердің дамуы туралы деректер”, Шванның 1839 ж. “Жануарлар мен өсімдіктердің Клетка теориясының негізгі қағидалары:
- барлық ұлпалар клеткалардан түзіледі;
- өсімдіктер мен жануарлар клеткаларының құрылысы ұқсас, өйткені барлық клеткалардың пайда болуы бір заңдылыққа бағынады;
- әрбір жасуша дербес, ал ағзаның тіршілік етуі жеке клеткалардың тіршілігінің жиынтығы салдарынан туады; құрылымы мен өсуіндегі сәйкестік туралы микроскоптық зерттеулер” деген атақты еңбектерінде жарық көрді. Осы жылдардан бастап клетка теориясының негізі қалыптасты. Ф. Энгельс клеткалық теорияны XIX ғасырдағы үш ұлы жаңалықтың (энергияның тұрақтылық заңы, клеткалық теория және Ч. Дарвиннің эволюциялық ілімі) бірі деп жоғары бағалады. Клеткалық теория шыққаннан кейін 20 жылдан соң неміс дәрігері Рудольф Вирхов (1859) клетка клеткадан ғана пайда болады деген корытынды жасап клеткалық теорияны одан әрі дамытты. Қазіргі кездегі клеткалық теорияның негізгі қағидалары мыналар: 1. Клетка — тіршіліктің ең нәзік әрі кішкене бірлігі (единицасы);2. Түрлі организмнің клеткалары құрылысы жағынан ұқсас;3. Клеткалар бөліну арқылы көбейеді;4. Көп клеткалы организмдер клеткалар мен олардың туындыларының жиынтығы.
2.Цитоплазманың химиялық құрамы, құрылысы, қызметі. Цитоплазма (гр. kytos — жасуша және гр. плазма — қалыптасқан) — ядроны қоршап жатқан жасуша бөлігі; қоймалжың (коллоидті) ерітінді. Клетка Цитоплазмасының сырты плазмолеммамен қапталған. Плазмолемма — Цитоплазманың ақуызды-билипидті қа-бықшасы, оның орт. қалыңд. 6 — 10 нм, құрамында ферменттер болады. Ол жасуша мен оны қоршаған орта арасындағы зат алмасу процесін қамтамасыз етеді. Цитоплазманың негізгі құрамы гиалоплазмадан, органеллалардан және қосындылардан тұрады.Тірі ағзалардың құрамына кіретін жасушаларды екі топқа бөлуге болады: ядро қабаты болмайтыны прокариоттар оларға көк жасыл балдырлар және бактериялар жатады, ядро қабаты жақсы жетілгені эукариоттар, бұған өсімдіктер мен жануарлар жасушалары жатады.Эукариотты жасушалар үш бөлімнен тұрады: сыртын қоршап жатқан плазматикалық мембранадан, цитоплазмадан және ядродан.Прокариоттар мен өсімдік жасушаларының плазматикалық мембраналарының сыртында жасуша қабаты болады, ал жануарлар жасушаларында мұндай қабат болмайды. Клетканың ядросын қоршап жатқан қоймалжың затты цитоплазма деп атайды Эукариотты жасушалар цитоплазмасының әртектілігі оның құрамында гиалоплазманың болуынан. Гиалоплазма мембраналы және мембранасыз компоненттерден тұрады. Мембраналы компоненттерге митохондриялар, пластидтер, эндоплазмалық тор, Гольджи аппараты, лизосомалар, ал мембранасыз компоненттерге центриольдар, рибосомдар, микротүтікшелер, микрофиламенттер жатады. Цитоплазманың құрамындағы жоғарыда аталған компоненттер бір-бірімен өте тығыз байланыста болады. Гиалоплазма — Цитоплазманың нағыз ішкі ортасын түзетін, оның негізгі плазмасы. Онда ақуыз молекулалары кешендерінің түзілуі мен ыдырау процестері үздіксіз жүріп жатады. Гиалоплазма (гр. hyalinos — мөлдір, шыны тәрізді; гр. plasmos — плазма) — жануарлар және өсімдік жасушалары цитоплазмасының органеллалар (тұрақты кездесетін құрылымдар - жасушаның тым ұсақ мүшелері) мен қосындылар (тұрақсыз құрылымдар) орналасатын біркелкі қоймалжың заты, жасуша цитоплазмасының негізгі заты. Гиалоплазма жасушаның ішкі ортасын құрайды. Гиалоплазманың құрамына негізінен глобулалы протеиндер кіреді. Олар жалпы жасуша протеиндерінің 20-25% құрайды. Гиалоплазмада қант, азоттық негіздер, амин қышқылдары, липидтер т.б. қосылыстар метаболизмінің ферменттері болады. Гиалоплазмадағы бос рибосомалар мен полисомаларда жасушаның өз керегіне пайдаланылатын протеиндер түзіледі. Органеллалар (органеллы); (organellae, гр. organon — құрал, мүше, ағза; гр. еllа — майда, кішірейту) — жасуша тіршілігі үшін әртүрлі қызмет атқаратын жасуша цитоплазмасындағы тұрақты құрылымдар. Адам мен жануарлар жасушаларында органеллалардың екі түрі болады. Олар: жалпы органеллалар және арнайы органеллалар. Жалпы органеллалар ағза жасушаларының барлығына түгелдей ортақ, яғни барлық жасушаларда кездеседі. Өз кезеңінде жалпы органеллалар: жарғақты (мембраналы) және жарғақсыз органеллалар болып екі топқа бөлінді. Жарғақты органеллаларға: митохондриялар, гранулалы эндоплазмалық тор, агранулалы (тегіс) эндоплазмалық тор, пластинкалық кешен (Гольджи кешені), лизосомалар, ал жарғақсыз органеллаларға: бос рибосомалар, полисомалар, центросома (центриольдер), микротүтікшелер, кірпікшелер, талшықтар және микрофиламенттер жатады. Арнайы органеллалар тек кейбір жасушаларда ғана болады. Оларға миофибриллалар (ет жасушаларында, ет талшықтарында — миосимпласттарда), тонофибриллалар (эпителиоциттерде), нейрофибриллалар (нейроциттерде), кірпікшелер, талшықтар жатады. Өсімдіктер жасушаларында аталған жалпы органеллалардан басқа пластидтер (хлоропласт, хромопласт, лейкопласт) болады. Органеллалар жалпы және арнайы болып бөлінеді. Жалпы органеллалар — жануарлар мен өсімдіктер жасушаларында болатын және тұрақты қызметтер атқаратын құрылымдар. Арнайы органеллалар — тек кейбір маманданған жасушаларға тән (биожіпше, тоножіпше, нейрожіпше, кірпікшелер, микробүрлер). Бұлардың әрқайсысының атқаратын қызметтері бар. Органелла жасушасының қаңқасын түзетін және оның митоздық бөлінуін қамтамасыз ететін органелла — центросоманы құрайды. Жоғары сатыдағы өсімдік жасушалары Гиалоплазмасында центросома центриольдері болмайды, оларда арнайы органоидтар — пластидтер (лейкопластар, хлоропластар, хромопластар) болады, ал Гольджи кешені Органеллада шашырап жатқан диктиосома түрінде берілген. Қосындылар — Органелланың тұрақсыз құрылымдары. Бұған: трофик. (ақуыздар, майлар, көмірсулар), секреторлық (биол. белсенді заттар), пигменттік (каротин, гемоглобин, билирубин, меланин, т.б.) заттар жатады. Өсімдік жасушасының Органелласында өсімдік шырынына толған, арнайы жарғақпен (тонопластпен) қапталған қосынды — вакуольдер болады; Цитоплазма мембраналары. Клеткада кездесетін барлық мембраналар (плазматикалық органоидтардың мембраналары. Мембраналардың құрылыстарын анықтайтын көптеген модельдер ұсынылды. Олар негізінде ақуыздар мен фосфолипидтердің байланысын көрсетеді. Кейбір модельде мембрана үзілмейтін қатпарлы құрылым, кейбіреуінде мицелла түрінде көрсетіледі. Мембрананың ішкі глобулярлы түйіршіктері ашылғаннан кейін мембраналарды ақуыздар кіретін сұйық құрылым ретінде қарайды және ішкі жасуша мембраналары құрылысы және химиялық құрамы жағынан бір-біріне ұқсас келеді. Электронды микроскоптан мембрананың үш қабаттан тұратынын анық көруге болады. Ортаңғы ашық қабат бимолекулалы май қабатынан, ал шеткі күңгірт қабаттары ақуыз молекулаларынан түзіледі. Клетканың сыртқы қабаты (плазматикалық) оны сыртқы ортадан бөліп тұрады, ал ішкі мембраналар көпіршік түрінде келіп, өзінің құрамындағы заттарды цитоплазмадан ажыратады. Клетка цитоплазмасындағы мембраналар қуыстар, түтікшелер немесе көпіршіктер жасап, өздерінің ішіндегі заттарды гиалоплазмадан бөліп, күрделі тор құрайды. Мұндай мембраналарға митохондрия, пластидтер, эндоплазмалық торлардың мембраналары жатады. Плазмалемма. Клетканың ішкі мембраналарына қарағанда плазмалемма холестерипке бай. Плазмалемманың бір ерекшелігі оның сыртында көмірсулардан тұратын гликокаликс қабаты орналасады. Бұл қабаттың қалындығы 3—4 нм-дей болады. Плазмалемма негізгі атқаратын қызметі: қорғаныштық, өткізгіштік және тасымалдаушы. Тасымалдаушы плазмалемма сулардың, иондардың және молекулалардың сыртқы ортадан жасушаға өтуін және кері өтуін реттеп отырады. Зат алмасу процесінде жасушада пайда болған қорытылған заттар да осы плазмалемма арқылы сыртқа шығарылып отырады. Плазмалемманың сыртқы бетіне рецепторлық ферменттер орналасады, олар жасушаның күйін басқа көрші жасушаларға жеткізіп тұрады. Плазмалемма жасушаның бөліну процесінде маңызды рөл атқарады. Оның сыртында микротүтікшелер, талшықтар сияқты әр түрлі өсінділер болады. Плазмалемманың тасымалдаушы қызметі. Үлкен молекулалар жай өтеді де, кіші молекулалар жылдам өтеді. Ең жылдам өтетін су және оның құрамындағы ерітінділер екен. Егер эритроцитті өзінің цитоплазмасынан гөрі шоғырлануы аз тұзды ерітіндіге салсақ, онда сыртқы ортадан су жасушаға көп енеді де жасушаның көлемі өсіп, оның сыртқы мембранасы жарылып кетеді. Керісінше, егер эритроцитті шоғырлануы көп тұзды суға салсақ, онда жасушадағы су бөлініп шығады да, ол қабысып, жиырылып қалады. Клетканың сыртқы мембранасында линопротеин қабатында «порлар» болады, олар арқылы иондар және сулар өтеді. Осы мембрана арқылы К+, N+ катиондары аса жылдамдықпен жүреді. Мембрананың тасымалдау қызметі: иондар мен судың шоғырлануы көп ортадан шоғырлануы аз ортаға (пассивті тасымалдау), және керісінше, шоғырлануы аз заттың Шоғырлануы көп ортаға енуін (активті тасымалдау) реттеп отырады. Активті тасымалдау энергия жұмсау арқылы өтеді (АТФ), К+, Na+ -иондарының шоғырлануын реттеп тұратын К+, Na+—«насосы» плазмалеммаға орналасады. Осы «насостардың» жұмыстарын зерттеу үшін эритроцитті алуға болады. Бұл жасушада К+, Na+ иондарының шоғырлануы қанның плазмасына қарағанда өзгеше болады (бірақ концентрациясының жиынтығы шамалас, жасушаның сыртында және ішінде изотониялы). Белоктар, нуклеин қышқылдары, майлар, ыдырап барып мономерлер түрінде ғана плазмалеммадан өте алады.
3.. Плазматикалық мембрана, ультрақұрылысы және физико-химиялық құрылымдары Плазмалемманың бір ерекшелігі оның сыртында көмірсулардан тұратын гликокаликс қабаты орналасады. Бұл қабаттың қалындығы 3—4 нм-дей болады. Плазмалемма негізгі атқаратын қызметі: қорғаныштық, өткізгіштік және тасымалдаушы. Тасымалдаушы плазмалемма сулардың, иондардың және молекулалардың сыртқы ортадан жасушаға өтуін және кері өтуін реттеп отырады. Зат алмасу процесінде жасушада пайда болған қорытылған заттар да осы плазмалемма арқылы сыртқа шығарылып отырады. Плазмалемманың сыртқы бетіне рецепторлық ферменттер орналасады, олар жасушаның күйін басқа көрші жасушаларға жеткізіп тұрады. Плазмалемма жасушаның бөліну процесінде маңызды рөл атқарады. Оның сыртында микротүтікшелер, талшықтар сияқты әр түрлі өсінділер болады.
Кейбір жағдайларда макромолекулалар немесе ірі түйіршіктер жасушаға эндоцитоз процесі арқылы өтуі мүмкін. Эндоцитозды фагоцитоз жәме пиноцитоз деп екі түрге бөлуге болады: фагоцитоз процесі дегеніміз жасушалардың ірі түйіршіктерді қабылдап цитоплазмасына өткізуі. Бұл процесті бірінші рет зерттеген орыс ғалымы — И. И. Мечников. Пиноцитоз процесінде жасуша цитоплазмасына ерітінділерді қабылдайды. Плазмалемма арқылы ішінде сұйық заттары бар көпіршіктердің жасушаға өтуін пиноцинтоз деп атаймыз. Эндоцитоздың бірінші кезеңінде (адсобция) энергия жұмсалмайды. Сырттан келген көпіршіктер плазмалемманы итеріп, ойыс жасап барып өтеді немесе плазмалемма өсінділері біртіндеп жасушаға енеді, жасушаға енген көпіршіктер плазмалеммадан жеке бөлініп барып орналасады.

• Плазматикалық мембрананың құрылысы
• 1. Белоктар және липидтер
• 2. Гликокаликс, полисахаридпен байланысы
Гликокаликс (glycocalyx, грек, glykys — тәтті, calyx — тостағанша) — барлық жасушалар плазмолеммаларымен тығыз байланысып, олардың сыртқы қабатын (орташа қалыңдығы 3-4 нм) құрайды. Гликокаликс — тарамдалған тым жіңішке күрделі көмірсулар — гликопротеидтер мен гликолипидтер жіпшелерінен тұрады. Гликокаликсте жасуша плазмолеммасының сыртындағы күрделі молекулалы протеиндерді, майларды, көмірсуларды қорытып, оларды өздерінің қарапайым мономерлеріне айналдыратын ферменттер болады.
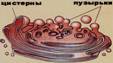
4. Гольджи кешенінің құрылысы мен қызметінің байланысын табыңыз. Гольджи кешені Алғашқы рет «Гольджи аппараты» деп аталатын органоидты 1898 жылы мишықтың Пуркинье клеткаларынан тауып ашқан италиялық ғалымы Камилло Гольджи. Гольджи аппаратын (кешенін) «ішкі тор тәрізді аппарат» деп те атайды. Бұл зерттеулердің бағасын көп жылдар өткеннен кейін ғалымдар қоғамы түсініп, оның авторына Нобель сыйлығы берілді. Гольджи аппаратының түрлі клеткаларда орналасуы әр уақытта ұқсас болып келеді. Көбінесе ол ядроның маңайында орналасып клетка орталығын қоршап тұрады. Кейбір клеткаларда Гольджи аппараты ядроның астында да орналасуы мүмкін. Электронды микроскоп арқылы зерттеулер бойынша Гольджи аппараты үш компоненттен тұрады.
1. Қатар орналасқан жазық мембраналардан тұратын жалпақ қапшықтар (цистерналар) жүйесі. Мұндай көршілес қатар жатқан қапшықтардың ара қашықтығы 14-15 нм. Әр қапшықтың ішкі қуысының ені 9 нм, ал мембраналардың қалыңдығы 7 нм.
2. Екінші компонент – белок секретін эндоплазмалық тордан тасымалдайтын диаметрі 30-50 нм болатын көпіршіктер.
3. Үшінші компонент – диаметрі 0,2-0,3 мкм болатын ірі вакуольдер.
Өсімдіктер және омыртқасыздардың клеткаларында Гольжди аппараты диффузды (батыраңқы) түрде клеткада жайылып жатады. Гольджи аппаратының мұндай түрін диктиосом деп атайды. Диктиосом жазық қапшықтардан (цистерналар) және олардың шеттерінде орналасқан майда көпіршіктерде тұрады. Алғашқы секрет цистерна мен майда көпіршіктерде қалыптасады, одан кейін ірі вакуольдерге түседі. Жетілген секреті бар ірі вакуольдер клетканың шетіне қарай қозғалады, олардың мембраналары плазмалеммамен қосылып кетеді де, секрет сыртқа шығады.
Гольджи аппаратының секрет шығарудағы қызметі радиоавтография әдісі бойынша таңбаланған атомдардың көмегімен ұйқы безінің экзокрин бөлігінде жан-жақты тексерілді. Жануарларға (3H - лейцин) амин қышқылды авторадиография әдісін қолданып клетка ішінде зерттелген. Бірінші таңбалар 3-5 минут өткеннен кейін түйіршікті эндоплазмалық торда табылған. Себебі, белок синтезі тек сол торда ғана жүреді. 20-40 минут өткеннен кейін таңба Гольджи аппаратының аймағында табылады, ал 60 минуттан кейін зимоген түйіршіктері де таңбаланады. Осындай тәжірибелер басқа клеткаларда да жасалған (сүт безінде, ішектің бокал тәрізді клеткасында, қалқанша безде т.б.).
Сонымен, рибосомдарда синтезделген белок түйіршікті эндоплазмалық тордың қапшықтарында жиналып Гольджи аппаратының аймағына жақындайды. Бұл жерде эндоплазмалық тордың тегіс шеттерінен құрамында синтезделген белогі бар майда вакуольдер бөлініп Гольджи аппаратының майда вакуольдерімен, немесе қапшықтарымен қосылады. Гольджи аппаратының бұл жағы проксимальді немесе цис полюсі деп аталады. Осылай синтезделген белок Гольджи аппаратының цистерналарына және ірі вакуольдерінде жиналып тығыздалады. Одан кейін секретті вакуольдер клетка мембранасына қарай жөнеліп плазмалеммамен қосылады. Сонда вакуольдің құрамындағы заттар сыртқа шығады. Бұл құбылысты экзоцитоз деп атайды. Гольджи аппаратының бұл жағы дистальді немесе транс полюсі деп аталады.
Вакуольдердің қозғалуын АТФ энергиясы және цитоплазманың микротүтікшелері мен микрофиламенттері қамтамасыз етеді.
Тағы да бір айтатын жайт синтезделген заттар гиалоплазмадан мембраналармен шектеліп тұрады. Бірақ Гольджи аппаратының қызметі тек секреттерді шектеу, жинау, тасымалдау, сыртқа шығаруда емес. Мұнда күрделі химиялық құбылыстар да өтеді. Тритиймен таңбаланған 3H – глюкозаны және 3H – галактозаны қолданып, ішек эпителиясының бокал тәрізді клеткалары зерттелген. Сонда 3H – глюкоза мен 3H – галактоза бірден Гольджи аймағында табылған. Осылай көмірсулардың Гольджи аппаратының цистерналарында синтезделетіндігі дәлелденген.
Өсімдіктің Гольджи аппаратында клетка қабығының матрикс (гелицеллюлоза, пектиндер) полисахаридтері синтезделеді. Ал клетка қабығының қаңқасы болатын целлюлоза арнайы ферменттер көмегімен плазматикалық мембрананың бетінде синтезделеді. Бірақ сол синтезге қажетті ферменттер Гольджи аппаратының вакуольді жүйе арқылы тасымалданады.
Сонымен бірге, Гольджи аппаратының вакуольдерінде липид молекуласы белоктарға қосылып – липопротеидтер жасалынады да сыртқа шығарылады.
Төменгі сатыдағы өсімдіктерде жиырылғыш вакуольдер Гольджи аппаратының цистернасынан пайда болады. Жиырылғыш вакуоль плазмалеммаға жақындап мембранасымен қосылғанда ішіндегі зат экзоцитоз жолымен сыртқа шығарылады.
Плазмалемманың жаңаруы да Гольджи аппаратына байланысты. Эндоплазмалық торда жасалған мембраналар Гольджи апаратының вакуольдеріне қосылып, Гольджи кешенінің аймағын ұлғайтады. Ал, Гольджи кешенінен мембраналық көпіршіктер плазмалеммаға қарай қозғалып, плазмалеммамен қосылып, оны жаңартып ұлғайтады. Гольджи аппаратының мембраналарының қалыңдауы оның гликокаликс (гликопротеид, гликолипид) қабатының пайда болуымен және жаңа белоктардың қосылуымен байланысты.
Лизосомдардың пайда болуы және атқаратын қызметі секреттік вакуольдерге ұқсас. Олар түйіршікті эндоплазмалық тор мен Гольджи аппаратының көмегімен пайда болады. Бірақ секреттік вакуольдер «экспортқа» жасалатын болса, клеткадан шығарылып басқа мүшелерде өз қызметін көрсететін болса, лизосомдар клетканың ішінде жұмыс істеп, эндогенді және экзогенді макромолекулаларды ыдыратады.
Сонымен Гольджи кешені клеткада өте маңызды қызметтер атқарады: 1) эндоплазмалық торда өндірілетін секреттерді қалыптастыру және сыртқа шығару; 2) көмірсулар синтезіне қатысу және көмірсуларды полипептид тізбектеріне жалғастыру; 3) лизосомдар құрастыруға қатысу; 4) сперматозоидта Гольджи аппаратының акросом қалыптасады.

5. Митоздық хромосомалардың жалпы құрылымы,түрлері және пішіндері. Әрбір ядрода хромосомалардың тұрақты саны және олардың әрқайсысының өзіне тән генетикалық мәні болады. Әрбір хромосома хромонема деп аталатын ДНҚ жіпшесінен тұрады, ол жіпшенің бойында тізілген моншақ сияқты құрылымдар – хромомерлер орналасқан. Хромосоманың міндетті түрде болатын құрылым ерекшелігіне бірінші бунақталу (перетяжка) жатады, ол хромосоманы екі бөлікке бөледі. Бірінші бунақталудың ішінде ерекше түзіліс – центромера болады, ол хромосомалардың митоздық таралуында маңызды роль атқарады. Центромера метофазадағы хромосомалардың пішінін анықтайды. Егер центромера хромосоманың ортасында орналасса және хромосоманы тең екі иыққа бөлсе, онда хромосоманың құрылымы Х - тәріздес болады, оны метацентірлі хромосома деп атайды. Центромера хромомсоманы тең екі иыққа бөлмесе, ондай хромосомаларды акроцентірлі хромосома деп атайды. Егер центромера хромосоманың ұшына жақын орналасса және хромосома иықтарының біреуі ғана анық көрінсе, телоцентрлі хромосома деп атайды. Кейбір хромосомалар ішінде екінші бунақталу болады. Кейде екінші бунақталудың көрінуі соншалықты анық, хромосома иықтарының біреуінің шеткі бөлігі ол хромосома мен тек жіңішке жіп арқылы ғана жалғанған сияқты болып көрінеді. Одай хромосома «спутникт і» (қосшы) хромосома деп аталынады. Клеткалар бөлінгенде хромосомаларың морфологиялық ерекшеліктері өзгермей сақталады. Клетка бөлінер алдында әр хромосома да бөлінеді.бұл процес ядродағы хромосомалар санының екі еселенуіне әкеледі. Сомалық клетка ядроларындағы ДНҚ-ның мөлшері жыныс клеткаларының ядроларына қарағанда екі есе көп болады. Клеткалардың бұл екі типі бір-бірінен хромосомалар саны арқылы ажыратылады. Жыныс клеткаларындағы хромосомалар саны гаплоидты деп аталады және «п» белгісі арқылы белгіленеді. Көп клеткалы организм денесінің сомалық клеткаларында хромосомалардың саны екі есе көп болады, және оны диплоидты деп атайды. Гомологты хромосомалардың әр жұбының біреуі аналық, екіншісі аталық организмнен келеді. Әр түрлі организмдердің сомалық тканьдері клеткаларындағы хромосомаларды зерттеу тканьдердің тек өзіне ғана тән хромосомасы болатынын көрсетті. Әр түрге тән хромосомалар жиынтығының ерекшеліктері – хромосомалардың саны, көлемі және формасы (пішіні) оның кариотипі деп аталады. Хромосомалар жұбымен топтастыра отырып, профазалық немесе метафазалық хромосомалардың микрофотографиялары бойынша идиограмма (хромосомаларды ұзындығы бойынша орналастыру) құруға болады. Дифференциалдық бояудың С- әдісі хромосома құрамындағы гетерохроматин бөлігін, яғни хромосомалардың басқа эухроматин бөліктеріне қарағанда, өте тығыз спиральға оралған хромосомаларды табуға мүмкіндік берді. Гетерохроматин констутивті және факультативті болып екіге бөлінеді. Мұның біріншісін хромосоманың белгілі учаскелерінен үнемі табуға болады, ал екіншісін клетка тіршілігінің белгілі бір кезеңдерінде ғана тек кейбір тканьдер клеткаларының хромосомасныда ғана көруге болады. Констутивті гетерохроматин үнемі гомологты хромосомаларлың ұқсас учаскелерінде болады, көпшілігнде центромераға жақын орналасады. Факультативті гетерохроматин – генетикалық активтіліктен толық айырылған хромосомалардың эухроматинді бөлігі. Мұндай хроматин гомологты хромосоманың бір сыңарында ғана болады. Кейбір жағдайларда жеке хромосомалар тұтасымен тек гетерохромтиндерден тұруы мүмкін.
6. Диффузды және тығыз хроматин (17срак)
Хроматин-ДНП болып келеді, ДНҚдан тұрады.Хроматиннің 2түрі белгілі. Ол эу-, гетерохроматин. Айырмашылықтары: эухроматин ядро ортасына жақын орналасқан, ашық түсті, деспирилизацияланған, аз тығыздалған, уникалды жұптардың қайталануы жоғары, С-фазада репликацияланады, мейозда рекомбинацияланады,белсенді гендері болады. Профазада ширатылып, қысқарады. Транскрипция жүретін ферменттерді активтендіреді. Гетерохроматин болса тығыз спиральданған, боялмайтын, сфаза соңында репликацияланады, ген болмағандықтан мейозда репликацияланбайды,теломера мен центромерада орналасқан, активті гендер анықталмаған бөлік б/т.. Оның 2түрі анықталған: констутивті (барлық клеткада гетерохроматин болып қала береді)сіңлілі хром-ң жабысуына әсер етеді;факультативті(кейде эухроматинге ұқсас болады, мыс. Инактивтелген Х-хром/сы немесе Барр денешігі. Ол барлық сүтқоректілердің аналығынан табылған, кариолеммада орналасады).
Репликацияда гетерохром эухром-нен кейін жатады.яғни гетерохр аймағы соңынан синтезделгендіктен жас өсіп келе жатқан клеткада анықталмаған.
Хроматин - жасуша ядросының негізгі компоненті; оны интерфазалық ядродан немесе митотикалық хромосомадан бөліп алу айтарлықгай жеңіл. Ол үшін оның әлсіз иондық күші бар сулы ерітіндіде немесе жай деиоидалғаи суда ерітілген күйге көшетін қасиетін пайдаланылады, алайда бірден с.йық күйге емес, қоймалжың күйге ауысады. Мұндай препараттарды нағыз сұйық ерітіндіге айналдыру үшін сілку, араластыру және қосымша гомогенизация сияқты күшті механикалық эсерлерді кажет етеді. Осындай әсерлер, сондай-ақ оны ұсак фрагменттерге бөлшектегенде хроматиннің қалыпты құрылымының бұзылуына әкеледі, бірақ оның химиялық құрамы өзгеріссіз қалады. Әр түрлі объектілерден алынған хроматин фракциялары айтарлықтай ұқсас немесе бірдей компоненттер жиынтығынан тұратындығы анықталды. Интерфазалық ядродан алынған хроматиннің химиялық құрамы митотикалық хромосоманың хроматинінен аз мөлшерде ерекшеленеді. Хроматиннің негізгі компоненттері ДНҚ және белок, ақуыздардың ішінде негізгісі гистон және гистондық емес белоктар. Бөлініп алынған хроматиннің орташа шамамен 40%-ға жуығын ДНҚ және 60%-ға ақуыздар құрайды, барлық белоктардың ішінде ядролық арнайы белок- гистондар 40-тан 80%-ға дейінгі мөлшерін құрайды. Хроматинді фракцияның құрамына одан басқа, РНҚ, көмірсулар, липидтер, гликопротеидтерсияқты мембраналы компоненттер де кіреді. Бірақ осы минорлы компонентердің хроматин құрылымына қанша мөлшерде кіретіндігі әлі анықталмаған. Тек РНҚ ДНҚ матрицасымен байланыстылығын жоғалтпаған күйіндс, яғни траскрипциялық РНҚ күйінде болуы мүмкін. Басқа да минорлы компонеіптср ядро кабықшасын құрайтын заттарға жатуы мүмкін.
Хроматиннің құрамында 30-40%-ы ДНҚ-ның еншісінде болады. Бұл ДНҚ, таза сулы ерітіндіден бөлініп алынған ДНҚ-ға ұқсас, қос спираль болып келеді. Бұл жайлы көптеген экспериментальді зерттеулер бар. Мысалы, хроматин ерітіндісін қыздырғандадаза ДНҚ-ны қыздырғандағыдай, ДНҚ-ның скі тізбегі арасындағы нулеотид аралық сутектік байланыс үзіліп, ерітіндіде гиперхромдық эффект деп аталатын жоғары оптикалық тығыздық қалыптасады. Хромосоманың құрылымын түтастай түсіну үшін хроматин құрамындағы ДНҚ молекуласы ұзындығының маңызы зор. Хроматин ДНҚ-сын бөліп алудың стандартты әдістерінде оның молекулалық массасы 7-9* 106 болады, Бұл ішек таяқшасы ДНҚ-ның молекулалық массасынан (2,8-109) әлдеқайда кіші. Хроматин препаратынан алынған ДНҚ-ның молекулалық массасының салыстырмалы түрде аз болуын, оны хроматин препаратынан бөліп алу процесі кезінде механикалық зақымдалулар эсерінен болуы мүмкін деп түсінуге болады. Егер ДНҚ-ны сілку арқылы, гомогенизация және басқа да жағдайлармен әсер ете отырып бөліп алса, онда ДНҚ молекуласын жасушадан өте үлкен ұзындықта алуға болады. Эукариотты жасушалардың ядросы мен хромосомасындағы ДНҚ молекуласының ұзындығын, прокариотты жасушаларды зерттегендегідей, фотооптикалық радиоавтография әдісінің көмегімен анықтауға болады. Хромосома құрамындағы жеке сызықтық ДНҚ молекуласының ұзындығы (прокариоттық хромосомалардан ерекшелігі) жүздеген микрометрден бірнеше сантиметрге жететіндігі анықталды. Әр түрлі объектілерден алынған ДНҚ молекуласының ұзындығы 0,5 мм-ден 2 см-ге дейін ауытқуы байқалды. Хромосомаға есептелінген ДНҚ-ның ұзындығы радиоавтографиялық әдіс арқылы алынған сандарға жақын келетінін осы нәтижелер көрсетіп отыр.
МИТОХОНДРИЯЛАР құрылысы қызметі
Митохондриялар (грекше митос — жіп және хондрион — түйіршік) — жіпше және түйіршік тәрізді органоид. Ол автотрофты және гетеротрофты организмдердің ци-топлазмасында кездеседі. Мнтохондрияларды ең бірінші 1850 жылы Р. А. Келликер насекомдардың бұлшық еттерінен байқады, оған «саркосома» деген термин берді (бұлшық еттегі митохондрияларды осы кезге дейін осылай атап жүр). Альтман (1890 жылы) арнаулы бояулар арқылы митохондриялардың анық көрінетінін дәлелдеп, оларды «биобластылар» деп атады. Бенде 1898 жылы бұл органоидқа митохондриялар деген ат берді. Михаэлис тірі клеткалардың митохондрияларын жасыл янус бояуымен бояп, олардың клеткадағы тотығу процестерімен байланысы бар екенін атап көрсетті.
Митохондриялардың көлемі тұрақты емес, сондықтан да олардың сыртқы пішіні әркез өзгермелі келеді. Көп клеткаларда олардың қалыңдығы тұрақты (0,5 мкм), ал ұзындығы тұрақсыз (жіпше тәрізді митохондриялар) 7—10 мкм-ге дейін жетеді. Митохондриялардың шын көлемін жарық микроскопымен анықтау қиын. Электронды микроскоппен митохондриялардың жұқа (400—500 А°) кесінділерін тексеру арқылы да оның көлемін дәлелдеу оңайға түспейді. Сондықтан да мүмкіндігінше митохондриядан алынған көптеген жұқа кесінділердің реконструкциясын (кеңістіктегі көлемі) жасап, оның нақты көлемін анықтауға болады.
Митохондрияның саны клетканың түріне қарай өзгермелі болады. Кейбір балдырлардың клеткаларында және қарапайымдарда бір ғана митохондрия, әр түрлі жануарлардың аталық жыныс клеткаларында (спермотозоид) олардың саны 20 — 70-ке дейін, сүтқоректілердің дене клеткаларында 500 — 1000-на дейін, ал алып амебада (Сһао8 сһаоз) 500000 дейін жетеді. Жануарлар клеткаларына қарағанда жасыл өсімдіктерде митохондриялар аз кездеседі, өйткені митохондриялардың кейбір қызметтерін хлоропластар атқарады.
Митохондриялар клетканың цитоплазмасында біркелкі, ал кей жағдайларда, әсіресе, патология кезінде, ядроның айналасына немесе цитоплазманың шет жағына қарай орналасады. Цитоплазмада клетка қосындылары (гликоген, май) көп болған жағдайда олар митохондрияларды клетканың шетіне ығыстырады. Митохондриялар митоз процесінде ұршық жіпшесінің айналасына шоғырланып, клетка бөлінгенде олар жас клеткаларға тең беріледі. Негізінде митохондриялар АТФ керек жерлерге миофибрилдерге тақау, ал сперматозоидтарда талшықты оран орналасады.
Сонымен митохондриялардың саны клетканың түріне және оның атқаратын кызметіне байланысты болады. Бауыр клеткасында болатын жалпы белоктың 30 — 35%-і митохондриялардың құрамында кездесетіні, ал бүйректе 20%-і болатыны анықталды.
Митохондрия екі мембранамен қоршалған, 6-7 нм шамасындай қалыңдығы бар, гиалоплазмадан бөліп тұратын сыртқы мембранадан және митохондрияның қүрылысына қарай күрделі өсінділер («криста») беретін ішкі мембранадан тұрады (14-сурет). Ішкі және сыртқы мембраналардың арасында ені 10-20 нм-ге тең кеңістік болады. Ішкі мембрана митохондрияның ішіндегі матрикс немесе митоплазмасын қоршап жатады. Күрделі өсінділерді немесе кристалар құратын мембраналардың ара қашықтыры 10 — 20 нм шамасындай болады. Кристалардың митохондрияларда орналасуы әр түрлі, кейбір клеткаларда көлденең бағытта орналасады, кейбіреулері тармақтанып келеді (15-сурет). Қарапайымдардың, бір клеткалы балдырлардың, кейбір жоғары сатыдары өсімдіктер мен жануарлардың клетка аралық митохондриялардың ішкі мембранасының өсінділері түтікше.
Митохондрияның құрамында белоктар (65—70% құрғақ салмағының) липидтер (25—30%), нуклеин кышқылдары (ДНҚ, РІНҚ) витаминдер және т. б. енеді. Митохондрияның құрамына енетін белоктардың көпшілігі — тотығу процесін қамтамасыз ететін, матриксінде және ішкі мембраналарына орналасқан ферменттер. Митохондриялардың қызметі осы ферменттерге байланысты.
Митохондрияның сыртқы мембранасының құрамындағы белоктар 20% болса, ал ішкі мембранасында 75%-ке дейін жетеді, мұның өзі оның басқа клеканың мембраналарына қарағанда ерекшелігін көрсетеді. Митохондрияның сыртқы мембранасы көрсеткіштері жағынан эндоплазмалық торға ұқсас. Сыртқы мембранада және мембрана аралық кеңістікте тотығу процесіне қатысатын ферменттер аз болады.
Сонымен митохондриядағы ферменттер клетканың тыныс алуына қажетті ферменттер болып табылады. Митохондрияның матриксінде «Кребс» цикліне қатысатын ферменттер шоғырланады. Ішкі мембранасында электрондарды тасымалдайтын тізбек және фосфорландыру процесіне қатысатын тасымалдау ферменттері (АДФ-тен АТФ) орналасады. Митохондрияда органикалық субстраттардың тотығуы және АДФ фосфорлануы нәтижесінде АТФ синтезделеді, сондықтан да митохондрияны клетканың күш беретін станциясы деп атайды. Клеткадағы тотығу және энергия жинау процестері бірнеше кезеңмен жүреді.
Бастапқы субстрат ретінде әр түрлі көмірсулар май қышқылдары, аминқышкылдары қолданылады. Көмірсулардың бастапқы тотығуы гиалоплазмада оттегісіз жүреді. Сондықтан оны анаэробты тотығу немесе гликолиз деп атайды. Анаэробты тотығудың негізгі субстраты — глюкоза. Кейбір бактериялар энергияны пентозаның, май қышқылдарының, амин қышқылдарының тотығуы арқылы алады. Бұл процесс мына теңдеуге сәйкес келеді:
 С6Н12O6 + 602 6Н2О + 6СО2 + 680 ккал.
С6Н12O6 + 602 6Н2О + 6СО2 + 680 ккал.
Клеткада энергия бірден бөлінбейді, ол сатыланып жүреді, химиялық энергия жылуға айналмайды, ол тек макроэнергиялық байланысқа АТФ-ке ауысады.
Гликолиз процесінде глюкоза триозаға дейін ыдырайды, мұнда 2 молекула АТФ жұмсалады да, 4 молекула АТФ синтезделеді, сонымен 1 моль глюкоза ыдырағанда 10% энергия жұмсалады. Гликолиз процесінде аз энергия жұмсалғанмен де бұл табиғатта жиі кездеседі. Микроорганизмдердің, кейбір ішек паразиттерінің, жаңадан дамып келе жатқан эмбриональды организмдердің клеткалары үшін гликолиз негізгі энергия көзі болып табылады. Сүтқоректілердің эритроциттері өздеріне керекті энергияны гликолиз арқылы алады, өйткені оларда митохондриялар болмайды. Гликолиз процесінде пайда болған триозалардың одан әрі тотығуы осы митохондриялардың өздерінде жүреді. Мұнда барлық химиялық қосылыстардан ыдыраған энергия қолданылады, осыған байланысты СО2 бөлінеді және оттегін қолдана отырып көп мөлшерде АТФ синтезделеді. Бұл процестер трикарбон кышқылының тоғыруымен жүреді. Осыдан АТФ-тың фосфорлануы арқылы АТФ молекулалары синтезделеді. Митохондрияларда толық белок синтездейтін жүйе болады, осыған байланысты ол өзінің ДНҚ-сы арқылы РНҚ молекулаларын синтездейді. Митохондрия құрамында рибосомдар болғандықтан, белок синтезі тұрақты жүреді. Митохондриялардың құрамындағы ДНҚ-ның ядродағы ДНҚ-дан айырмашылығы болады (молекулалық салмағы жағынан және нуклеотидтердің құрамы және орналасуы жағынан).Митохондрияда жүретін ДНҚ синтезінің ядродағы ДНҚ синтезімен байланысы жоқ, олар өз ферменттері арқылы ғана байланысады. Митохондриялардың матриксында ДНҚ матрицасы арқылы РНҚ синтезі өтеді. Митохондрияда РНҚ-ның информациялық, тасымалдаушы, рибосомды түрлері синтезделеді.
Қызметі:
Митохондриялар АТФ-ті синтездеуші органелла. Негізгі функциясы қосылыстарды толықтырып, олардың ыдырауының нәтижесінде бөлінген энергияны пайдалануға байланысты. Осы қызметін еске алып, Клод митохондрияларды клетканың «қуат станциясы» деп атаған. Клеткалық тыныстың органелласы деп атауға да болады.
Клетка ішіндегі сұйық затты сіңіру және бөліп шығаруда митохондрияның маңызы үлкен, катиондарды жинау қызметін де атқарады.
Не нашли, что искали? Воспользуйтесь поиском: